фото, советы опытных декораторов по расстановке мебели в разных комнатах
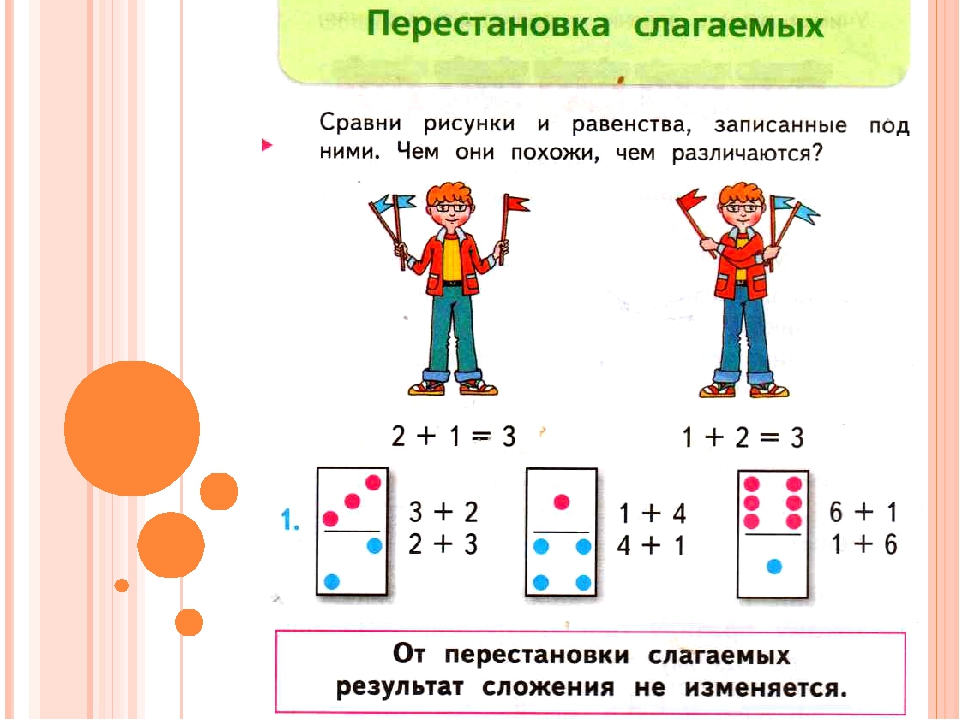
Владельцев крошечных квартир и домов волнует, как расставить мебель в маленькой комнате? Небольшими комнатами считаются от 6 кв.м. до 20 кв.м. Комната в 20 кв.м. может показаться солидной, но если это однокомнатная, или трехкомнатная квартира, а спальня, детская, гостиная по 8 кв.м., тогда маленькие пространства могут быть очень функциональны и комфортны, если знать, как обустроить маленькую комнату. В статье мы разберем, как сделать комнату уютной: переставить мебель и найти дополнительные места для хранения вещей и освобождения пространства.
Оглавление статьи:
Расставляем мебель – основные принципы
Перед тем как переставить мебель в небольшой квартире, нужно определиться какие комнаты самые маленькие. Возможно, получиться их разгрузить, чтобы оставить только необходимый набор предметов и расширить пространство.
Для разгрузки комнаты отыскивают незадействованные места.
Также помещение может стать тесным, если в ней находятся нужные, но редко используемые предметы обстановки. Их можно заменить складывающимися моделями. Например, стол-трансформер можно просто сложить и убрать до прихода гостей.
После этого начинается расстановка мебели в маленькой комнате. Существует 3 основных варианта того, как расставить мебель:
- симметрично;
- асимметрично;
- по кругу.
Симметричная обстановка выполняется вокруг одного предмета, взятого за центр. Например, вокруг дивана или телевизора. Остальная мебель должна расположиться на равном отдалении. Этот вариант подходит для классического дизайна, с единым цветом и размером элементов.
Асимметричная обстановка чаще всего применяется в маленьких комнатах. Для неё нет определенных требований как в симметричном способе. Но каждый элемент должен быть в одной стилистике. Среди мебели и аксессуаров разных размеров и цветом, нужно отыскать подходящие, чтобы не получить захламления.
Для неё нет определенных требований как в симметричном способе. Но каждый элемент должен быть в одной стилистике. Среди мебели и аксессуаров разных размеров и цветом, нужно отыскать подходящие, чтобы не получить захламления.
Чтобы расставить мебель по кругу, нужно также как в симметричном, определиться с центром. После всё распределяется на равной удаленности, получая правильный круг. Это сложнейшая расстановка, потому что нельзя смещать предметы.
Для гармоничной обстановки меленьких комнат нужно пользоваться четырьмя принципами.
Практичный подход
Подбирая для маленькой комнаты мебель, стоит задуматься, для чего именно она необходима. Возможно, это лишний элемент, который займет и так ограниченное место. Для маленьких пространств чаще применяются трансформируемые столы, стулья и др. Для получения дополнительного места для хранения вещей, диван подбирается с ячейками и боковыми полками. Для спален кровать должна быть со стеллажами в изголовье и выдвигающимися ящичками.
Максимальная разгрузка
Для получения максимальной функциональности перестановка мебели в маленькой квартире должна заполнить незадействованные места. Например, углы комнаты. В некоторые можно установить диван, чтобы было больше места для расположения гостей. Над ними можно повесить открытые полочки, чтобы сложить накопившиеся книги.
Вместо дивана можно установить узкие стеллажи для дополнительного хранения вещей или сделать «домашний офис» с маленьким столом, ноутбуком и светильником.
Разгружают комнаты системами хранения под окнами и над ними. Над рамами устанавливается открытая полка с коробками, а можно заказать навесные шкафы. Если шкафы над окном будут закрытые, то фасад должен быть из прозрачной пластмассы или глянцевый, чтобы не отяготить обстановку.
Вертикальное хранение
Многих волнует вопрос, как можно обставить маленькую комнату, чтобы увеличить свободное пространство. С этим справляются прикроватные тумбы, компактные стеллажи, этажерки различной формы на колесиках и без них.
С этим справляются прикроватные тумбы, компактные стеллажи, этажерки различной формы на колесиках и без них.
Функциональное использование подоконника
Для более функционального использования пространства перестановка некоторой мебели в комнате может быть на подоконник. Для кухни он может стать обеденной или рабочей зоной. В гостиной можно организовать книжный уголок, «домашний офис». А в спальне туалетным столиком.
Расстановка мебели в комнатах разного назначения
Гостиная
Перед расстановкой мебели в гостиной нужно определить количество людей, которые будут в комнате и их вид досуга. У молодой семьи должно быть большое свободное место. Можно установить барную стойку и интересное освещение, чтобы получить расслабляющую обстановку для вечернего времяпровождения.
Семейные пары чаще применяют круговую остановку: в середине ставится маленький столик, а вокруг кресло, диван. Большая семья нуждается в просторном помещении. Для экономии места мебель встраивают или приобретают не громоздкую и трансформируемую. Если зал очень узкий, то можно всё расставить у стенок друг напротив друга. Например, диван напротив телевизора и открытых полок.
Если зал очень узкий, то можно всё расставить у стенок друг напротив друга. Например, диван напротив телевизора и открытых полок.
Интересно! Если комната слишком мала, то окно и подоконник оставляют свободными, чтобы попадало больше света.
Если шкаф купе можно разместить только в зале, то подбирают с зеркальными фасадами для визуального увеличения пространства.
Кухня
Кухня — это одна из самых посещаемых мест в доме. Она должна быть функциональной и уютной. Планировка маленькой кухоньки может быть линейной, Г-образной, двухрядной.
Линейно располагается гарнитур и столовая зона в самых маленьких пространствах: вдоль одной стены устанавливается кухонный гарнитур со столом-трансформером в конце в качестве обеденной зоны или обеденное место переносится в другую более просторную комнату.
Для Г-образного расположения устанавливается угловой кухонный гарнитур, а обеденный стол ставят напротив или переносят в другое место.
В двухрядном шкафчики гарнитура устанавливаются друг напротив друга или шкафы ставятся с одной стороны, а параллельно располагают барную стойку или откидной обеденный стол.
Интересно! Комфортным расстоянием от кухонного гарнитура до обеденной зоны 90-120 см. Поэтому в крошечных кухнях не советуют устанавливать обеденный стол по центру.
Спальня
В спальне очень важно сохранить уют, комфорт и расслабляющую атмосферу. Мебель лучше подбирать теплых и светлых оттенков.
Центром комнаты является кровать. Для крошечных квартир были придуманы модели с маленьким изголовьем. Если помещение очень узкое, кровать устанавливают возле маленькой стены. Также шкаф можно подобрать до потолка и поставить вдоль короткой стенки — помещение станет похожа на квадрат. В комнатах в форме квадрата кровать устанавливается изголовьем в стенке, а тумбочки с обеих сторон. Туалетный столик ставится у окошка. Для спален в форме прямоугольника, кровать ставят к стене удлиненной стороной.
Для малогабаритных квартир устанавливают встроенные или модульные шкафы. Комод в крохотных комнатах не ставится, его можно заменить открытыми полками или выдвижными шкафами шкафа. Также можно вместо стандартного спального места, установить диван-тахту с подъемным механизмом. У него нет спинок и подлокотников, что позволяем трансформироваться в полноценную кровать. К тому же там имеется много места для постельного белья.
Интересно! Если рабочее место находится в спальне, то его можно установить на подоконнике и разделить зоны ширмой.
Детская
Детская комната должна быть стильной, ведь вкусы формируются с детства. А также функциональной, экологичной и безопасной. Кроватка должна быть с высокими бортиками, это обезопасит малыша во время сна. В настоящее время много кроваток, которые трансформируются по мере взросления малыша.
Если в комнате спят два ребенка, то устанавливается кровать с двумя ярусами. Чтобы дети чувствовали защищенность, она ставится у стены подальше от окон. Для маленьких детей нужен небольшой стол с ящиками, чтобы складывать карандаши, раскраски и т.п. Школьникам нужен полноценный стол ученика с ящиками для канцелярии, тетрадок и книг. Если устанавливается компьютер, то его нужно ставить на компьютерный стол с множеством ящиков и выдвигающейся полкой для мышки, клавиатуры.
Для маленьких детей нужен небольшой стол с ящиками, чтобы складывать карандаши, раскраски и т.п. Школьникам нужен полноценный стол ученика с ящиками для канцелярии, тетрадок и книг. Если устанавливается компьютер, то его нужно ставить на компьютерный стол с множеством ящиков и выдвигающейся полкой для мышки, клавиатуры.
Интересно! Компьютерные столы устанавливают возле окна, чтобы ребенок не портил зрение от недостатка света.
Полезные рекомендации
Избежать типовых ошибок можно, если знать их «в лицо» и следовать рекомендациям:
- Перестановка мебели без предварительного замера. В малогабаритных квартирах всё хочется расположить как можно плотнее. Но лишние бездумные перестановки только повредят мебель, пол и стены. Перед процедурами измерьте всё рулеткой или сантиметром.
- Перестановка без учета всех мнений в доме. Например, если вы передвинули рабочую зону без учета мнения домочадца, который находится там каждый день – недопонимание неизбежно.
 Возможно, там для него слишком плохое освещение и это повредит здоровью. Поэтому учитывайте мнения всех.
Возможно, там для него слишком плохое освещение и это повредит здоровью. Поэтому учитывайте мнения всех. - Иногда после передвижения мебели чувство удовлетворенности не наступает. Возможно, вам лучше избавиться от лишних предметов и заметить их новыми? Например, надоевший ковер или кресла с некрасивой обивкой.
- Перемещение только крупногабаритной мебели. Создают уютную атмосферу именно мелочи: тумбочки, столики, светильники, картины. Поэтому не забывайте их тоже переставлять вместе с крупными предметами.
- При кардинальной перемене функциональных зон, необходимо изменение освещения. Оно должно соответствовать, чтобы поддерживать ощущения комфорта.
- Передвигание мебели не должно нарушать принятых пропорций, чтобы не получить нефункциональную обстановку. Стандартные пропорции изучают заранее. Например, что между двух диванов должно быть не меньше 1м. и 10см., а в прихожей должно быть не менее 0,5м. свободного места.
- Переставлять мебель привычными, негармоничными способом. Например, если у вас всегда возле кресла стояла массивная тумба и снова переносите их в новое место комплектом, то никакого преображения не произойдет. К тому же собирая в одной стороне комнаты крупные предметы, а другой мелкие – потеря баланса очевидна.
 Возможно, там для него слишком плохое освещение и это повредит здоровью. Поэтому учитывайте мнения всех.
Возможно, там для него слишком плохое освещение и это повредит здоровью. Поэтому учитывайте мнения всех. Например, если у вас всегда возле кресла стояла массивная тумба и снова переносите их в новое место комплектом, то никакого преображения не произойдет. К тому же собирая в одной стороне комнаты крупные предметы, а другой мелкие – потеря баланса очевидна.
Например, если у вас всегда возле кресла стояла массивная тумба и снова переносите их в новое место комплектом, то никакого преображения не произойдет. К тому же собирая в одной стороне комнаты крупные предметы, а другой мелкие – потеря баланса очевидна.Фото-подборка идей расстановки мебели в маленькой комнате
Посмотрите фото идеи по расстановке мебели для маленькой комнаты:
Борис Джонсон провел существенные перестановки в британском кабмине и заменил главу МИД — Международная панорама
ЛОНДОН, 15 сентября. /ТАСС/. Премьер-министр Великобритании Борис Джонсон провел серьезные перестановки в своем кабинете, поменяв ряд ключевых министров. В отставку или на другие должности были отправлены несколько членов кабинета, в том числе и те, действия которых последнее время активно критиковались.
По мнению аналитиков, таким образом Джонсон преследует две цели — снизить недовольство однопартийцев своим кабинетом и объединить Консервативную партию, уже задумываясь о выборах 2024 года.
«Сформированный мной сегодня кабинет будет без устали работать над объединением страны и повышением уровня жизни на всей ее территории. Наша цель после пандемии — восстановить все лучше, чем было, и выполнить поставленные приоритетные задачи. Возьмемся же за работу», — написал Джонсон на своей странице в Twitter, повторяя лозунг Консервативной партии Build back better, что будет звучать все чаще в преддверии выборов, которые могут состояться и раньше намеченного срока.
Самым громким стал переход министра иностранных дел Доминика Рааба в министерство юстиции. Хотя Рааб и остался заместителем главы правительства и даже подкрепил этот статус новой формальной должностью, его аппаратный вес на новом посту будет значительно меньше, чем во главе Форин оффиса, которым он руководил с 2019 года.
На смену Раабу пришла министр внешней торговли, по делам женщин и равноправия Элизабет Трасс. Таким образом, из четырех главных министерских портфелей в Великобритании (МИД, МВД, Минфин и Минобороны) два принадлежат женщинам. Ранее стало известно, что Прити Пател, которой многие обозреватели прочили отставку, сохранила за собой пост министра внутренних дел. В руководстве Минфина и Минобороны изменений проведено не было.
Ранее стало известно, что Прити Пател, которой многие обозреватели прочили отставку, сохранила за собой пост министра внутренних дел. В руководстве Минфина и Минобороны изменений проведено не было.
Почему сместили Рааба
Британский политолог, редактор и издатель журнала Politics First Маркус Пападопулос назвал причиной смещения Рааба с поста главы МИД тот факт, что министр находился в отпуске, когда члены радикального движения «Талибан» (запрещено в РФ) захватывали Афганистан. В итоге, по оценке телеканала Sky News, эвакуация британцев была проведена не столь эффективно, как это сделали другие страны, например, Франция. Хотя Лондон сумел вывезти из Кабула около 15 тыс. гражданских лиц, власти королевства признали, что в Афганистане остались сотни британцев, а также многие из помогавших им на протяжении 20 лет иностранной военной операции афганцы.
«Он очень медленно отреагировал на изменение ситуации в Афганистане, и на то, как быстро «Талибан» завоевал страну и пришел ко власти. В глазах многих людей, не только в парламенте, правительстве, но и обычных британцев, его репутация сильно пострадала. Поэтому для Бориса Джонсона было важно убрать его с этой должности», — сказал Пападопулос.
В глазах многих людей, не только в парламенте, правительстве, но и обычных британцев, его репутация сильно пострадала. Поэтому для Бориса Джонсона было важно убрать его с этой должности», — сказал Пападопулос.
Другие назначения
В рамках перестановок в британском кабмине своих постов лишились министр образования Гэвин Уильямсон и министр жилищного строительства, по делам регионов и местного самоуправления Роберт Дженрик, которые, по мнению обозревателей, не справлялись с возложенными на них обязанностями. Роберта Бакленда, чей портфель достался Раабу, напротив, называют исключительно компетентным министром юстиции, однако новой должности в кабинете для него не нашлось.
Дженрика сменил канцлер герцогства Ланкастерского (министр по особым поручениям) Майкл Гоув, которого считают способным справиться с жилищным кризисом в Великобритании. Оставленную Гоувом должность занял Стив Баркли, который в 2018-2020 годах занимал пост министра по делам Brexit. Экс-министр по делам международного развития Энн-Мари Тревелиан получила принадлежавший Трасс пост министра внешней торговли, однако новая руководительница МИД продолжит отвечать за вопросы равноправия и права женщин. Уильямсона на посту главы Минобраза сменил замминистра по делам бизнеса, энергетики и промышленной стратегии королевства Надим Захави, который отвечал за программу вакцинации от коронавируса и очень хорошо себя зарекомендовал.
Уильямсона на посту главы Минобраза сменил замминистра по делам бизнеса, энергетики и промышленной стратегии королевства Надим Захави, который отвечал за программу вакцинации от коронавируса и очень хорошо себя зарекомендовал.
Министр цифровых технологий, культуры, средств массовой информации и спорта Великобритании Оливер Дауден передал свой портфель Надин Доррис, занимавшей пост заместителя министра здравоохранения, и получил пост председателем Консервативной партии, на котором будет заниматься вопросами партийной организации. Именно на них он специализировался в правительстве Дэвида Кэмерона (2010-2016). При этом Дауден останется в правительстве в качестве министра без портфеля. На должность министра без портфеля в секретариат кабмина также был назначен Найджел Адамс, ранее отвечавший за азиатское направление в Форин оффисе.
Зачем были нужны перестановки
Пападопулос считает, что Джонсон преследовал как минимум две цели, затеяв масштабные перестановки, всего лишь вторые с момента прихода к власти.
«За последние полтора года Великобритания прошла через очень сложный этап, связанный с пандемией COVID-19. На многих членов кабмина обрушился шквал критики, причем не столько со стороны СМИ, сколько от собственных однопартийцев, не входящих в правительство. Комитет 1922, объединяющий рядовых депутатов, так называемых заднескамеечников, обладает возможностью отправить с отставку лидера партии. Именно он сыграл важную роль в отставке Терезы Мэй в 2019 году. Точно так же сейчас он заявил Джонсону, что в кабинете министров нужны новые лица», — сказал он.
Вторая причина, по мнению политолога, заключается в том, что Джонсон уже задумывается о будущих выборах.
«Они должны состояться через три года, но, возможно, что пройдут и раньше — уже через два. Действия премьера во время пандемии уже были предметом критики Комитета 1922. Вероятно, Джонсон хочет оставить все разногласия позади и объединить партию перед выборами», — заметил Пападопулос.
21. Перестановка в расписании.
 Рукописный девичий рассказ
Рукописный девичий рассказ21. Перестановка в расписании
Сегодня т. е. 31 ноября заболела Вера Васильевна и за отсутствием преподавателя истории, 10 [-м] классам переставили уроки.
Первым был урок алгебры.
Наша учительница, желая отдохнуть от нас, дала нам «маленькую» самостоятельную. Так как в нашем классе чувство товарищества выражено в большой степени, мы быстро с ней справились, и сейчас занимались кто чем. Тут всё и началось. В дверь постучали. Наша Валентина Павловна вышла. Побыв за дверью не больше минуты, она снова зашла в класс, но не одна: с ней была красивая, стройная девушка. Ее красота поразила всех. В нашей классной комнате создалась такая тишина, что было слышно, как тикают часы на руках. Юноши украдкой вздохнули, потому что поняли, что покой их отныне исчез: в незнакомку нельзя было не влюбиться. Девушки рассматривали ее одежду и волосы, которые были действительно красивы: длинные черные косы, чуть вьющаяся челка. На секунду все забыли, для чего она здесь.
Тут Валентина Павловна сказала: «Ее зовут Олеся. Она будет учиться в нашем классе месяц. Олеся! Ты потом им расскажешь о себе, а пока садись…»
— Сюда!
— У меня свободно!
— Я хочу с ней сидеть!
Раздались голоса с разных парт.
— Садись с…
Но Олеся улыбнулась и подошла к парте Стаса Друбича и села с ним. Стас самый симпатичный парень в нашем классе. Но он у нас недавно.
Приехал из Саратова к тетке, познакомился с ребятами и решил остаться. О себе рассказывать не любил. И мало кто знал о нём что-нибудь. Девушкам он нравился, но ему не нравилась ни одна из них, хотя в нашем классе есть красивые девушки. Стас хорошо учился, был серьезным человеком и здорово рисовал.
Звонок. Урок алгебры закончен. Все уходят в физ. кабинет. На перемене все расспрашивают Олесю.
— Мы живём в Саратове. А сюда не надолго, — объясняет она.
Только Стас вышел из класса. Зашел он лишь после звонка. На физике Олеся села снова с ним.
Весь урок они сидели молча, глядя на доску, и лишь в конце Стас посмотрел на нее. Красивые, тонкие черты лица: изогнутые дугами черные брови, длинные ресницы; прямой не большой нос; коралловые губы с родинкой под ними.
Олеся повернула голову — тяжелые косы упали вперед. Движением головы она откинула назад челку. Стас улыбнулся. Ее удивительные большие глаза внимательно посмотрели на Стаса, она улыбнулась.
Последние два урока прошли быстро. Прозвенел звонок с последнего. Все ушли домой, в классе остались лишь Стас и Олеся. Стас встал, подошел к окну. Олеся подошла к нему и сказала:
— Что ж, здравствуй, Стас…
— Здравствуй, Леся! Давно же мы не виделись с тобой. Кажется, полгода… Длинных и жестоких…
— Как же жил эти пол года, Стас?
— Плохо[,] Леся! Ведь ты была далеко… А ты?
Олеся молча смотрела Стасу в глаза.
— Когда ты уехал, — начала она, — я думала, что все будет хорошо. Даже радовалась, дурочка, что ты уехал. Но поняла, что не могу без тебя. Спасибо тебе за открытки, Стас. Спасибо, что не забыл…
— И никогда не забуду! Леся милая, я звал тебя! Я люблю тебя! Я не могу без тебя жить!
Глаза Олеси наполнились слезами.
— Стас… я приехала… я приехала к тебе… я хочу тебе сказать.., что я люблю тебя, Стас!
По лицу Леси катились слезы. Стас вытирал их.
— Стас! Забудь все, что я тогда сказала, прости меня, милый!
— Олеся! Любимая! Не плач[ь,] не надо. Давай забудем нашу ссору, начнем все сначала.
— Да, как тогда, помнишь, когда ты впервые сказал мне, что любишь…
Он обнял ее, Олеся нежно гладила его волосы.
— Леся! Пойдем[,] я покажу тебе твой портрет. На нем ты улыбаешься. Ты больше никогда не будешь плакать, ты будешь смеяться, как всегда. Пойдем!..
Когда они вышли из школы, было уже темно. Белый пушистый снег заметал их следы, а вскоре и совсем скрыл от посторонних глаз державшихся за руки Лесю и Стаса. Не верите, посмотрите расписание 31[-го ноября] действительно переставлено.
Рассказ получен от Л. Уразаевой, 1980 г. р., г. Шадринск, 2000 г.
Песков объяснил перестановки в правительстве — РБК
В Кремле прокомментировали отставку Цуканова и назначение Якушева, а также возможное повышение Новака. Вопрос, продолжит ли последний в качестве вице-премьера курировать работу с ОПЕК, Песков посоветовал адресовать правительству
Фото: Екатерина Штукина / ТАСС
В Кремле перестановки в правительстве назвали «ротационным вопросом», передает корреспондент РБК.
«Я не могу ответить на этот вопрос. Это ротационный вопрос», — сказал пресс-секретарь президента Дмитрий Песков.
Ранее Владимир Путин отправил в отставку полномочного представителя президента в Уральском федеральном округе Николая Цуканова, а на освободившееся место полпреда УрФО назначил бывшего министра строительства Якушева.
Также свободным стал пост министра энергетики Александра Новака. Его кандидатуру премьер-министр Михаил Мишустин выдвинул на должность вице-премьера.
Отвечая на вопрос, останется ли Новак курировать работу с ОПЕК и ОПЕК+, Песков посоветовал обратиться к правительству. «Подождите. Произойдет назначение, утверждение Думой, потом будем говорить», — отметил пресс-секретарь.
кадровые перестановки в правительстве Японии
С. Абэ рассчитывает на то, что смена кабинета создаст новый эффект ожиданий, который ранее помогал правительству обрести поддержку в массах
В начале августа премьер-министр Японии С. Абэ провел кадровые перестановки в кабинете министров. Столкнувшись с сильной критикой извне, С. Абэ в своих кадровых решениях предпочел действовать крайне осторожно и не прибегать к резким шагам, чреватым для него существенными политическими рисками. В новом составе кабинета министров преобладают неяркие политики умеренных взглядов из числа опытных и доказавших свою личную преданность С. Абэ ветеранов. По этой причине новый кабинет вызывает сильное чувство дежавю.
Новый выход на политический небосклон уже достаточно известных и относительно популярных деятелей второго эшелона призвано, с одной стороны, апеллировать к ностальгическим чувствам японцев, желающих вернуть то время, когда при власти С. Абэ у них «все было хорошо», с другой — отмыть правительство от неприятного налета, оставленного недавними политическими скандалами.
Главная задача для премьера заключается в том, чтобы восстановить рейтинги доверия правительству и удержать в течение года вплоть до выборов председателя ЛДП. Однако не имея широкой народной поддержки, С. Абэ вряд ли сможет рассчитывать на переизбрание. В ближайшем будущем японский премьер, скорее всего, бросит все свои силы на решение первоочередных вопросов экономического и социального развития. С большой долей вероятности японский премьер отодвинет на задний план задачу изменения конституции.
В начале августа премьер-министр Японии С. Абэ провел кадровые перестановки в кабинете министров. Решение отправить в отставку правительство было принято на фоне катастрофического падения рейтингов, связанного с целой чередой скандалов вокруг самого С. Абэ и членов его кабинета.
Некоторые из этих скандалов уже привели к отставкам отдельных членов кабинета. Только в 2017 г. на компромате с внебрачными связями, опубликованном японскими СМИ, «погорел» заместитель министра экономики торговли и промышленности Т. Накагава. Был вынужден уйти из правительства после высказываний, крайне сомнительных с моральной точки зрения, министр по делам ликвидации последствий фукусимской аварии М. Имамура. Уже в июле в отставку подала министр обороны Т. Инада, взяв на себя ответственность за сокрытие министерством обороны информации о деятельности миротворческого контингента Сил самообороны в Южном Судане, где жизнь японских военнослужащих подвергалась опасности (такая информация должна стать публичной, так как по действующей конституции персонал Сил самообороны не может быть задействован в тех точках, где не действует режима прекращения огня).
Несколько скандалов затронули и самого С. Абэ. Сначала его имя стало фигурировать в связи с сомнительной с юридической точки зрения земельной сделкой, которая принесла большую выгоду ультранационалистическому образовательному фонду Моритомо гакуэн, пользовавшемуся публичной поддержкой японского премьера. Затем один из друзей С. Абэ, руководитель фонда Какэ гакуэн, получил от главы кабинета личную протекцию в деле регистрации ветеринарной академии на территории особой экономической зоны. С. Абэ обвинили в фаворитизме и использовании служебного положения, а оппозиция развернула активную кампанию за его отставку.
Череда шумных разбирательств привела к тому, что поддержка правительства снизилась летом 2017 г. до отметки ниже 30% (по опросу газеты «Майнити» за 22–23 июля — до 26%). Критичность ситуации для правящей Либерально-демократической партии становится особенно очевидной, если учесть, что на протяжении более четырех лет с момента возвращения С. Абэ во власть в декабре 2012 г. рейтинги кабинета не падали ниже отметки в 50%. Ущерб репутации С. Абэ достиг такой степени, что брожение пошло даже внутри правящей партии, которая до последнего времени находилась под его авторитарным контролем.
Вариантов реагирования на создавшуюся ситуацию у премьера было не так много. Он мог ничего не делать, игнорируя критику в свой адрес, — и тогда рисковал утратить остатки общественной поддержки, поставив под удар само правление ЛДП. Другая возможность — роспуск парламента с целью получить новый кредит доверия со стороны избирателей — оказалась неприемлемой по причине непредсказуемости результатов внеочередных всеобщих парламентских выборов. Даже в случае победы С. Абэ рисковал утратить имеющееся сейчас у правящей коалиции в обеих палатах квалифицированное большинство, позволяющее ему провести через парламент поправки конституции. К тому же С. Абэ ставит перед собой задачу пойти в 2018 г. на третий срок на посту председателя ЛДП и остаться у руля правления страной в период проведения Токийской олимпиады 2020 г., которую он считает своим личным проектом. Ослабление позиций ЛДП в парламенте в результате внеочередных выборов поставило бы достижимость этой цели под вопрос, поэтому не хотел бы иметь никаких политических «неожиданностей».
Поэтому оставался единственный вариант — провести кадровую перестановку правительства. Согласно японской политической традиции, ответственность за грехи старого кабинета министров «не пристает» к новому правительству, даже если кое-кто из предыдущего состава сохраняет в нем свои посты.
Новый старый состав
Что можно сказать о новом составе правительства? Прежде всего, обращает на себя внимание явная осторожность японского премьера в выборе кандидатов на министерские посты. Из 19 членов кабинета только шесть парламентариев, совсем не имеющих за своими плечами министерского опыта, — в составе всех четырех кабинетов министров с момента прихода С. Абэ к власти в 2012 г. это наименьшее их число. С. Абэ опирался в первую очередь на свою «старую гвардию», которая хорошо зарекомендовала себя в удачный для него период первых четырех лет правления. Например, посты в правительстве сохранили политические «тяжеловесы» в лице вице-премьера и министра финансов Т. Асо и генерального секретаря кабинета министров Ё. Суга. Остались в новом составе кабинета на прежних должностях также министр экономики торговли и промышленности Х. Сэко (специальный министр по делам экономического сотрудничества с Россией), министр по делам ликвидации последствий катастрофы М. Ёсино и министр государственных земель и инфраструктуры К. Исии. Некоторые члены правительства остались в его составе, получив новые назначения (например, министр по делам экономического оживления Т. Мотэги и министр труда и здравоохранения К. Като).
Кстати, в ходе кадровой перетряски, затронувшей не только правительственную, но и партийную номенклатуру, ключевые должности в ЛДП оставили за собой и ее высшие руководители — генеральный секретарь ЛДП Т. Никаи и заместитель председателя ЛДП М. Комура.
Другой момент, достойный пристального внимания, — это возвращение в состав кабинета ряда политиков из числа членов предыдущих кабинетов С. Абэ, правивших страной начиная с 2012 г. Новый выход на политический небосклон уже достаточно известных и относительно популярных деятелей второго эшелона призвано, с одной стороны, апеллировать к ностальгическим чувствам японцев, желающих вернуть то время, когда при власти С. Абэ у них «все было хорошо», с другой — отмыть правительство от неприятного налета, оставленного недавними политическими скандалами.
В числе таких «новых старичков» особенно выделяется министр обороны Ицунори Онодэра, сменивший проштрафившуюся в результате упомянутого выше «миротворческого» скандала Т. Инаду. Онодэра уже возглавлял министерство обороны в 2012–2014 гг., а в последние годы прославился активной позицией по вопросу об перевооружении Сил самообороны. Группа депутатов под его руководством инициировала программу принятия на вооружение Сил самообороны крылатых ракет, которые относятся к наступательному оружию, а также модернизированных систем ПРО Aegis Ashore.
Другой знаковый политик подобного рода — новый министр образования и науки Ё. Хаяси, который своим приходом должен «отмыть» министерство образования от грязи, принесенной скандалами с образовательными фондами Моримото гакуэн и Какэ гакуэн. По словам помощника С. Абэ, одним из главных мотивов премьера стало «обеспечение стабильности и завершение склок в министерстве образования».
Еще одно важнейшее кадровое решение касается назначения на пост министра иностранных дел. С. Абэ выбрал Таро Коно, который в 2015–2016 гг. уже входил в правительство в качестве главы ведомства по делам административной реформы и председателя комиссии по общественной безопасности.
Политическая династия Т. Коно хорошо известна в Японии. Отец нынешнего главы дипломатического ведомства Ёхэй Коно, возглавлявший в середине 1990-х гг. ЛДП, получил большую известность в связи со сделанным им в 1993 г. заявлением, в котором он признал ответственность Японии перед т.н. женщинами для утешения — кореянками, пострадавшими от сексуального насилия со стороны военнослужащими японской императорской армии в годы Второй мировой войны. В последние годы Ё. Коно неоднократно выступал с критикой политики С. Абэ в области национальной безопасности.
Однако взгляды Т. Коно на проблемы обороны и национальной безопасности все же ближе к действующему премьеру, чем к отцу. На пресс-конференции после вступления в должность он подтвердил свою приверженность курсу на укрепление военных и политических отношений с США, участие Японии в международных соглашениях о свободной торговле и борьбу с потеплением климата. В то же время Т. Коно проявляет некоторую самостоятельность и даже либерализм в своих суждениях, улавливая общественные настроения, которые не всегда соответствуют «генеральной линии» партии. Так, в противоположность официальной позиции ЛДП он неоднократно выступал за немедленное закрытие всех атомных станций; за пересмотр соглашения о статусе американских вооруженных сил в Японии; за более открытую иммиграционную политику. В прошлом Т. Коно даже высказывался в поддержку идеи о юридическом признании для японцев двойного гражданства.
Несколько особняком среди прочих членов нового правительства стоит фигура нового министра внутренних дел и коммуникаций Сэйко Нода. Наряду с сохранившей свой пост в новом кабинете министром юстиции Ё. Камикава она стала одной из двух женщин в составе нового кабинета. В сферу ответственности С. Нода входят вопросы реформы почт и телекоммуникаций, являющиеся сферой ее специализации, и на новом поприще она, как ожидается, сможет в полной мере проявить свои политические способности.
Необычность назначения С. Нода заключается в том, что она относится к числу открытых внутрипартийных противников С. Абэ, представляя т.н. «антиосновное течение» партии. В 2015 г., когда истек первый срок С. Абэ, она выступила с резкой критикой японского премьера за его согласие провести фактически безальтернативные выборы председателя ЛДП и даже заявила о намерении побороться с ним. По всей видимости, включение С. Ноды в состав правительства призвано продемонстрировать, что С. Абэ, которого часто упрекают за авторитарный стиль правления, готов выслушивать своих политических противников и работать с ними в одной команде.
Межфракционный расклад сил
Еще один существенный аспект переформирования кабинета министров касается фракционной политики правящей партии. Многих обозревателей интересует, в какой степени С. Абэ принимает во внимание межфракционный расклад сил внутри партии и продолжает ли он традиции кадрового строительства на принципах фракционной политики, которые существенно пошатнулись после правления Дз. Коидзуми в 2001–2005 гг.
Можно заключить, что баланс сил между фракциями был соблюден — все они, за исключением фракции Исихары, получили как минимум по одному посту в правительстве. Однако обращает на себя внимание тот факт, что наибольшее количество министерских постов — четыре — было получено фракцией Фумио Кисиды (в прошлом кабинете их было два). Поскольку речь идет об относительно немногочисленной, четвертой по численности фракции ЛДП, появляются основания предположить, что С. Абэ при принятии кадровых решений особенно выделяет политические интересы Ф. Кисиды. В этой связи нелишне заметить, что наряду с Сигэру Исибой Ф. Кисида в настоящий момент рассматривается как наиболее сильный политический конкурент С. Абэ, способный успешно побороться за высший партийный пост. Примечательно, что в результате кадровых пертурбаций Ф. Кисида переместился с поста министра иностранных дел на чисто партийную должность председателя Совета по политическим вопросам ЛДП. Дистанцировавшись от правительства после пяти лет работы в нем, Ф. Кисида, таким образом, облегчает себе задачу номинировать свою кандидатуру на выборах председателя ЛДП, намеченных на сентябрь 2018 г. Знаменательно также, что не самая многочисленная в ЛДП фракция С. Исибы, одного из наиболее активных оппонентов С. Абэ внутри партии, получила три поста в правительственной иерархии (К. Сайто, Х. Кадзияма и Х. Оконоги). Скорее всего, премьер пытается подобным образом балансировать между двумя своими наиболее вероятными преемниками, не показывая явного предпочтения никому из них. Это позволяет сохранять интригу в вопросе о преемничестве и тем самым усиливать позиции самого С. Абэ.
Эффект дежавю
Столкнувшись с сильной критикой извне, С. Абэ в своих кадровых решениях предпочел действовать крайне осторожно и не прибегать к резким шагам, чреватым для него существенными политическими рисками. В новом составе кабинета министров преобладают неяркие политики умеренных взглядов из числа опытных и доказавших свою личную преданность С. Абэ ветеранов. По этой причине новый кабинет вызывает сильное чувство дежавю. На какой-то период это несколько повысит рейтинги власти, так как японцы в своей массе консервативны, не желают резких перемен и склонны с ностальгией вспоминать первые относительно благополучные годы курса «абэномики», а потому надеются на повторение «чуда» 2012–2014 гг., когда экономическое положение в стране действительно улучшалось. Именно «абэномика» на протяжении нескольких лет поддерживала популярность кабинетов С. Абэ, хотя многие из принимавшихся законов были крайне непопулярными — например, законы о защите государственной тайны или законы об обеспечении национальной безопасности. С. Абэ рассчитывает на то, что смена кабинета создаст новый эффект ожиданий, который ранее помогал правительству обрести поддержку в массах.
В ближайшем будущем японский премьер, скорее всего, бросит все свои силы на решение первоочередных вопросов экономического и социального развития. Уже на первой пресс-конференции после инаугурации нового кабинета С. Абэ заявил о намерении «запустить полезный круг экономического развития и покончить с дефляцией». Одновременно японский премьер с большой долей вероятности отодвинет на задний план задачу изменения конституции, которую он неоднократно декларировал в качестве первоочередной и даже обозначал конкретный срок ее реализации. Конституционный вопрос расколол японское общество, и премьер не собирается рисковать, продавливая его решение силовыми методами. С. Абэ уже заявил, что собирается отдать приоритет восстановлению экономического роста, а не пересмотру конституции.
Главная задача для премьера заключается в том, чтобы восстановить рейтинги доверия правительству и удержать в течение года вплоть до выборов председателя ЛДП. Однако не имея широкой народной поддержки, С. Абэ вряд ли сможет рассчитывать на переизбрание.
Получится ли С. Абэ повернуть вспять процесс утраты ЛДП кредита доверия среди избирателей? На первый взгляд, в определенной степени ему это удается. По итогам опросов общественного мнения, проведенным газетой «Майнити» вскоре после формирования нового кабинета, уровень поддержки правительства вырос на 9% по сравнению с июлем, составив 35%, тогда как уровень неодобрения составил 47%, снизившись на 9 пунктов. Однако цифры эти не должны никого обманывать. Станет ли восстановление доверия к правительству и лично к С. Абэ долгосрочным и необратимым, будет зависеть от успехов японской экономики.
В Московском музыкальном театре проведена перестановка в руководстве балета
Андрей УваровОткрыв новый сезон, Московский академический Музыкальный театр имени Станиславского и Немировича-Данченко (МАМТ) произвел перестановку в руководстве своей балетной труппы.
Возглавлявший балет МАМТ народный артист РФ Игорь Зеленский с 1 сентября 2016 года приступил к исполнению обязанностей интенданта и художественного руководителя Баварского государственного балета (Германия, Мюнхен).
В МАМТ ему предложено остаться в качестве советника гендиректора по балету. Исполняющим обязанности художественного руководителя балета МАМТ назначен управляющий балетной труппой, народный артист РФ Андрей Уваров.
Об этих кадровых перестановках сообщил генеральный директор МАМТ Антон Гетьман.
Предложение гендиректора“О том, что Игорь Зеленский с 2016 года станет интендантом Баварского государственного балета, было известно прежнему руководству МАМТ, которое считало, что он сумеет совмещать работу в Мюнхене и в Москве.
Но за месяц плотной работы в Мюнхене Игорь Анатольевич понял, что не сможет уделять театру Станиславского и Немировича-Данченко необходимое количество времени, и принял решение полностью посвятить себя работе в Баварском балете”,
– сказал Гетьман.
“Однако весь наступивший сезон сверстан именно Игорем Зеленским, а потому я предложил Игорю Анатольевичу на переходный период остаться моим советником по балету, чтобы иметь возможность совместно решать художественные и производственные вопросы. С 1 октября Игорь Зеленский – советник гендиректора МАМТ по балету”,
– сказал Гетьман.
Развивая тему сотрудничества, гендиректор театра упомянул о гастролях балета МАМТ в Мюнхене, которые состоятся в апреле 2017 года.
Позиция ЗеленскогоИгорь Зеленский принял предложение гендиректора МАМТ остаться его советником по балету и специально прилетел из Мюнхена в Москву, чтобы закрепить отношения с МАМТ в новом качестве.
“Я принял предложение Антона Александровича Гетьмана”,
– подтвердил Зеленский. Он назвал это предложение “очень приятным для себя”.
“Считаю, что в качестве советника по балету смогу и впредь быть полезен Театру Станиславского и Немировича-Данченко, в котором на посту худрука балетной труппы прослужил пять лет”,
– заключил Зеленский.
Игорь Зеленский – солист балета, народный артист РФ. Окончил Тбилисское хореографическое училище, стажировался в Ленинградском хореографическом училище имени Вагановой. В 1988 году принят в труппу Мариинского театра, где быстро достиг положения премьера.
Был также приглашенным солистом Большого театра, Берлинской оперы и Королевского балета Великобритании, премьером в New York City Ballet. В качестве главного приглашенного солиста танцевал в Ла Скала, Баварском государственном балете, театрах Бостона, Сан-Франциско, Рио-де-Жанейро.
В 2006 году стал художественным руководителем балетной труппы Новосибирского государственного академического театра оперы и балета. В 2011 году возглавил балет Московского академического Музыкального театра имени Станиславского и Немировича-Данченко, совмещая работу в Москве и Новосибирске. В 2016 году истек срок контракта Зеленского в Новосибирском театре.
Новая роль Андрея УвароваСо своей стороны, исполняющий обязанности худрука балета МАМТ, народный артист РФ Андрей Уваров поблагодарил за оказанное доверие.
“Новую должность я расцениваю не только как признание своей работы в качестве управляющего балетной труппой, но и как стимул для дальнейшей деятельности на новом посту”,
– сказал Уваров.
Андрей Уваров – народный артист РФ. Выпускник Московского хореографического училища, по окончании которого был принят в балетную труппу Большого театра, где стал ведущим солистом.
Завершив артистическую карьеру в 2011 году, Уваров был приглашен в качестве педагога-репетитора в МАМТ, где вскоре занял пост управляющего балетной труппой.
Ольга Свистунова, ТАСС
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Перестановка
Перегруппировки, вызванные катионными или электронодефицитными сайтами |
|---|
Катионные перегруппировки
В первой половине девятнадцатого века считалось, что реакции органических соединений протекают с минимальными структурными изменениями. Этот принцип упростил объяснение многочисленных реакций замещения, присоединения и элиминирования, которые характеризовали поведение общих функциональных групп.Однако последующие открытия показали, что природа не всегда была такой услужливой, оставляя химиков и студентов-химиков бороться с возможностью глубоких структурных изменений, происходящих во время определенных реакций. Большое количество этих структурных перестроек запускается промежуточными продуктами, включающими положительно заряженные или электронодефицитные атомы, которые в случае углерода являются карбокатионами. Двумя такими примерами, как уже отмечалось, являются добавление HCl к 3,3-диметил-1-бутену и принудительный гидролиз неопентилбромида.В этой главе будут описаны и обсуждены другие случаи этой интригующей группы преобразований.
1. Перестановки Вагнера-Меервейна
Химическое поведение неопентилбромида, 2,2-диметил-1-бромпропана, является поучительным местом для начала этого обсуждения. Очень низкая реакционная способность этого 1º-бромида S N 2 была отмечена ранее и объяснена стерическими препятствиями для требуемого 180º выравнивания реагирующих орбиталей. В условиях, благоприятствующих реакционной способности S N 1, таких как раствор во влажной муравьиной кислоте, неопентилбромид реагирует примерно с той же скоростью, что и этилбромид.Оба эти соединения представляют собой 1º-алкилгалогениды, и для реакции S N 1 на стадии определения скорости требуется ионизация до 1º-карбокатиона. Как отмечено в приведенном ниже порядке стабильности карбокатионов, такие карбокатионы относительно нестабильны и образуются медленно. Продуктом этилбромида является этанол, продукт простого и прямого замещения, но неопентилбромид дает 2-метил-2-бутанол вместо ожидаемого неопентилового спирта. Совершенно очевидно, что произошло изменение того, как пять атомов углерода в этом продукте связаны друг с другом.
|
Образовавшийся этильный катион может быть преобразован только в процессе замещения или отщепления.В случае неопентильного катиона, однако, первоначально образованный 1º-карбокатион может быть преобразован в более стабильный 3º-карбокатион за счет 1,2-сдвига соседней метильной группы с ее связывающими электронами. Механизм, демонстрирующий такую перестройку, показан ниже, и он очень хорошо объясняет общие структурные изменения.
Повышение стабильности промежуточных продуктов карбокатиона — не единственный фактор, который приводит к молекулярной перегруппировке. Если угловая деформация, скручивающая деформация или стерическое скопление в структуре реагента могут быть уменьшены за счет алкильного или арильного сдвига к сайту карбокатиона, такая перегруппировка обычно наблюдается.Следующие ниже примеры иллюстрируют перестройки, вызванные деформацией в маленьком кольце. Хотя первоначально образуется 3º-карбокатион, угол и деформация кручения четырехчленного кольца уменьшаются за счет сдвига метиленовой группы, что приводит к расширению кольца до 2º-карбокатиона. При нажатии на диаграмму уравнения отобразится механизм этих преобразований.
После стадии расширения кольца могут иметь место другие реакции, в зависимости от условий. В водной кислоте перегруппированный 2º-карбокатион может связываться с водным нуклеофилом, образуя 2º-спирт, терять протон в воду, давая 3,3-диметилциклопентен (не показан), или подвергаться второй перегруппировке в 3º-карбокатион, который затем образует 1,2-диметилциклопентен.Действительно, нередко можно встретить последовательности перегруппировок в более сложных соединениях, и они могут давать продукты со структурой, заметно отличающейся от структуры исходного соединения. Следующее уравнение показывает одну из таких реакций. Изогнутая стрелка, изображающая пять последовательных шагов расширения кольца, будет добавлена к уравнению, если щелкнуть диаграмму.
В терминологии перициклических реакций 1,2-алкильные сдвиги такого рода классифицируются как [1,2] -сигматропные сдвиги .Поскольку это двухэлектронный процесс (2 электрона в перемещенной сигма-связи), предполагается, что перегруппировка будет надлицевой. Как показывает следующий пример, были получены значительные доказательства, подтверждающие этот вывод. Протонирование двойной связи дает 3º-карбокатион. Соседний атом водорода (окрашенный в синий цвет) сдвигается как гидридная составляющая, чтобы создать новый 3º-карбокатион, который, в свою очередь, вызывает сдвиг метильной группы (окрашен в зеленый цвет) с образованием еще одного 3º-карбокатиона.Этот электрофильный центр затем связывается с нуклеофильным кислородом функциональной карбоновой кислоты, высвобождая каталитический протон для продолжения процесса. Из-за слитой полициклической структуры этого соединения легко определяется относительная ориентация мигрирующих групп, и видно, что она является супрафациальной. Перестройки, состоящие из последовательных 1,2-сдвигов, часто происходят согласованным и, следовательно, стереоспецифическим образом; однако не следует предполагать, что групповые смены происходят одновременно.Каждый сдвиг включает в себя отдельное переходное состояние, в котором положительный заряд делокализован по конечной точке миграции, исходной точке и мигрирующей группе.
Многие из наиболее интересных перегруппировок такого рода были обнаружены в ходе структурных исследований природных соединений. Среди них терпены представляли множество замечательных реакций, и имена двух химиков, которые сыграли важную роль в раскрытии их сложных превращений, Х. Меервейна и Г.Вагнера, постоянно связаны с этими перестановками. Добавление газообразной HCl к α-пинену оказалось особенно загадочным для этих первых химиков. В обычных условиях этот жидкий компонент скипидара давал кристаллическое соединение C 10 H 17 Cl, первоначально называвшееся «искусственной камфарой», теперь известное как борнилхлорид. Нестабильный изомер, гидрохлорид пинена, можно выделить в мягких условиях, но он быстро изомеризуется в борнилхлорид. Обработка борнилхлорида основанием дала кристаллический изомер пинена, называемый камфен, вместе с небольшими количествами другого ненасыщенного углеводорода (борнилена).Добавление HCl к камфену аналогичным образом первоначально дает нестабильный хлорид (гидрохлорид камфена), который быстро изомеризуется в изоборнилхлорид, стереоизомер борнилхлорида. Теперь мы знаем, что борнилхлорид и изоборнилхлорид являются эндо / экзо-2-хлор изомерами 1,7,7-триметилбицикло [2.2.1] бициклогептановой системы. Структурные формулы этих соединений представлены ниже вместе с камфеном, перегруппированным продуктом отщепления.
Механизмы этих перестановок будут изображены при нажатии на диаграмму выше.На новом изображении мы видим, что и пинен, и камфен образуют 3º-карбокатионы, когда двойная связь протонирована. Перегруппировка в 2º-карбокатион способствует снятию напряжения небольших колец в случае пинена и облегчению стерического застоя в случае камфена. Однако это чрезмерное упрощение, которое игнорирует тот факт, что эти реакции происходят в неполярных растворителях, и вряд ли будут включать дискретные, несвязанные карбокатионы. Некоторые из стереоэлектронных эффектов, влияющих на эти реакции, будут показаны при повторном нажатии на диаграмму выше.Слева показаны структуры изначально образовавшихся нестабильных гидрохлоридов пинена и камфена. Оптимальное перекрытие орбиталей разрыва и образования связей требует приближения сдвигающейся алкильной группы с тыльной стороны к месту уходящего хлорид-аниона аналогично реакции S N 2. Хлорид-анион расположен на одной стороне карбокатиона, образованного алкильным сдвигом, и сразу же связывается с этой стороной трехкоординатного углерода. С этой точки зрения этих перегруппировок хлорид-анион никогда не избегает притягательного влияния своего катионного партнера, и стереоселективность продукта понятна.Кислотные катализаторы Льюиса (например, FeCl 3 ) катализируют эти перегруппировки, и продукт, предпочтительный при равновесии, представляет собой борнилхлорид.
Перегруппировка, которая происходит в условиях отщепления, катализируемого основанием, отражает затененную конфигурацию двухуглеродного мостика, несущего атом хлора. Из-за этой конфигурации не может быть достигнута антикомпланарная структура, благоприятствующая переходному состоянию E2. Синтезирование дает небольшое количество борнилена, но преобладает перегруппировка в предшественник камфена.Повторное нажатие на приведенную выше диаграмму будет переключать дисплеи.
Еще один пример замечательных перегруппировок, катализируемых кислотой, которые происходят с терпенами, наблюдался при исследовании сесквитерпенового кариофиллена (из гвоздичного масла). Здесь очевидно, что реактивные центры могут взаимодействовать и образовывать связи от одной стороны кольца среднего размера к другой стороне. Механизмы многих таких перегруппировок изучались и до сих пор изучаются с большим интересом.
2.Перегруппировка Пинакола
Пинакольная перегруппировка была первой молекулярной перегруппировкой, идентифицированной как таковая ранними химиками. Определяющий пример перегруппировки пинакола показан на следующей диаграмме. Сам пинакол производится восстановлением ацетона магнием, вероятно, посредством промежуточного кетила. Поскольку диол является симметричным, протонирование и потеря воды происходит с равной вероятностью по каждой гидроксильной группе. Полученный 3º-карбокатион относительно стабилен, и было показано, что он возвращается в пинакол в результате реакции в присутствии воды, меченной изотопами.Сдвиг 1,2-метила создает еще более стабильный карбокатион, в котором заряд делокализован за счет резонанса гетероатомов. Действительно, этот новый катион представляет собой просто сопряженную кислоту кетона пинаколона, который является продуктом повторяющихся перегруппировок, катализируемых переносом протона. Каждая стадия этой перегруппировки потенциально обратима, что продемонстрировано катализируемой кислотой дегидратации пинаколона (и пинакола) до 2,3-диметил-1,3-бутадиена в жестких условиях.
При анализе течения пинакольной перестройки необходимо учитывать множество факторов.К ним относятся:
• Какая гидроксильная группа теряется в виде воды? или Какой промежуточный карбокатион более стабилен?
• Какова внутренняя тенденция к смещению (миграционная способность) различных групп заместителей?
• Какое влияние на перегруппировку оказывают стерические затруднения и другие факторы деформации?
• Образуются ли эпоксиды как промежуточные соединения в пинакольной перегруппировке?
• Влияет ли стабильность продукта на результат конкурирующих реорганизаций?
• Выполните условия реакции (т.е.е. тип кислоты, концентрация, растворитель и температура) влияют на ход перегруппировки?
Было показано, что практически все эти факторы важны в одном или нескольких случаях, и полный анализ их сложного взаимодействия выходит за рамки этого текста. Тем не менее, несколько примеров будут представлены, чтобы продемонстрировать общий характер этого преобразования и проиллюстрировать действие некоторых из вышеупомянутых факторов. В первой реакции, показанной ниже, мы видим пример кинетического и термодинамического контроля продукта.При обработке слабой кислотой диол быстро перестраивается в альдегид путем сдвига 1,2-водорода в первоначально образовавшийся дифенил-3º-карбокатион. Более энергичная кислотная обработка диола или альдегида дает более стабильный фенилкетон (конъюгация фенильной и карбонильной групп). Механизмы этой и других реакций будут представлены при нажатии на диаграмму. Стрелка розового цвета обозначает перестановку; светло-голубые стрелки указывают на реакции замыкания или размыкания эпоксидного цикла.Повторное нажатие переключает реакцию и отображение механизма.
Во втором примере описывается аналогичная реагирующая система, которая предоставляет дополнительную информацию из стереохимических и изотопных характеристик мечения. Потеря воды из сайта 3º-карбинола с последующим обратимым 1,2-гидридным сдвигом приводит к образованию конъюгированной кислоты кетонового продукта. При коротких временах реакции рацемизация регенерированного исходного диольного материала происходит с той же скоростью, что и перегруппировка. Соответствующий фенильный сдвиг к первоначально образованному 3º-карбокатиону генерирует сопряженную кислоту с альдегидом, и было показано, что сам альдегид изомеризуется в такой же перегруппированный кетон в условиях этой перегруппировки пинакола.Изотопная углеродная метка (окрашенная в зеленый цвет) в диоле или альдегиде перемешивается (окрашивается в коричневый цвет) в ходе этих реакций, что указывает на промежуточное соединение эпоксида.
В реакции № 3 в качестве реагента можно использовать цис- или трансдиол. Эти изомеры быстро взаимопревращаются в условиях перегруппировки, указывая на то, что начальная потеря воды обратима; результат подтвержден изотопным кислородным обменом. Явное предпочтение сдвигу метиленовой группы по сравнению со сдвигом метильной группы может отражать присущие им миграционные способности или, возможно, групповые конфигурации в промежуточном 3º-карбокатионном промежуточном продукте.В показанной здесь конформации могут сдвигаться как метильная, так и метиленовая группы, или может образовываться эпоксидное кольцо обратимо. Альтернативная конформация в виде кресла, имеющая экваториальную метильную группу, должна быть более стабильной, но не подходящей для метильного сдвига. Таким образом, понятно преобладающее сокращение кольца. Реакция №4 — необычный случай, когда напряженное кольцо сжимается до еще меньшего кольца. Фенильные группы обычно обладают высокой способностью к миграции, поэтому невозможность получить 2,2-дифенилциклобутанон в качестве продукта может показаться неожиданным.Однако карбокатион, возникающий в результате фенильного сдвига, будет таким же напряженным, как и его предшественник; тогда как сдвиг кольцевой метиленовой группы генерирует ненапряженный катион, стабилизированный фенильными и кислородными заместителями. Конъюгативная стабилизация фенилкетона и отсутствие гибридизированных атомов углерода sp 2 в малом кольце также могут способствовать стабильности наблюдаемого продукта.
Наконец, реакция № 5 ясно показывает влияние условий реакции на состав продукта, но сложно объяснить, каким образом различные условия влияют на результат.Обработка холодной серной кислотой должна давать более стабильный дифенил-3º-карбокатион, и тогда смещение метильной группы приведет к наблюдаемому продукту. С другой стороны, действие кислоты Льюиса в уксусном ангидриде может избирательно ацетилировать менее затрудненный диметилкарбинол. В этом случае ацетат становится предпочтительной уходящей группой (предположительно координированной с кислотой), за которой следует 1,2-фенильный сдвиг. Сообщается, что симметрично замещенный изомерный диол (показан в заштрихованной рамке) перегруппировывается исключительно за счет метильного сдвига, но конфигурация исходного материала не указана (возможны два диастереомера).
Из-за влияния других факторов (см. Выше) невозможно определить однозначный порядок миграции заместителей в перегруппировке пинакола. Однако можно выделить некоторые общие тенденции. Бензопинакол, (C 6 H 5 ) 2 C (OH) C (OH) (C 6 H 5 ) 2 , подвергается быстрой перегруппировке в (C 6 H 5 ) 3 CCOC 6 H 5 в гораздо более мягких условиях, чем требуется для пинакола.Действительно, часто бывает так, что фенил или другие ароматические заместители, соседние с образующимся карбокатионом, будут способствовать этой ионизации в ходе их миграции к катионному сайту. В неароматических соединениях типа (CH 3 ) 2 C (OH) C (OH) RCH 3 миграция R возрастает следующим образом: R = CH 3 2H 5 3) 3 ° C. Поскольку замещающая алкильная группа должна нести часть общего положительного заряда, алкильное замещение должно оказывать стабилизирующее влияние на переходное состояние перегруппировки.Наконец, замещение фтора, как в случае C 6 H 5 (CF 3 ) C (OH) C (OH) (CF 3 ) C 6 H 5 , делает диол неактивным под действием кислотного катализатора. условия перестановки. Здесь мощный индукционный отвод электронов фтором препятствует образованию положительного заряда.
3. Перестановка Тиффно-Демьянова
Неопределенность в определении начального места образования карбокатиона представляет проблему при анализе многих перегруппировок пинакола.Эта неопределенность может быть устранена путем дезаминирования азотистой кислотой соответствующих 1º-аминоспиртов, как показано в следующем уравнении. Поскольку эту реакцию обычно проводят в очень мягких условиях, вероятность того, что последующие превращения могут скрыть начальную перегруппировку, значительно снижается.
Перегруппировка Тиффно-Демьянова часто используется для превращения циклического кетона в гомолог, который на один размер кольца больше. Такое применение, которое осуществляется посредством промежуточного циангидрина, показано в первом примере ниже.Циклические кетоны имеют два альфа-углеродных атома, каждый из которых может переходить в формирующийся 1º-карбокатион. Если в показанном здесь случае R = H, эти две группы идентичны и при сдвиге дают одинаковый продукт. Если R = CH 3 , 2º-алкильная группа предпочтительно смещается, причем основным продуктом является 3-метилциклогексанон; 2-метильный изомер является второстепенным продуктом. Вторая реакция информативна, поскольку демонстрирует, что хиральная 2º-бутильная группа перемещается с сохранением конфигурации.
Третий пример иллюстрирует важность конфигурации подложки в процессе перегруппировки.Начальную стадию сдвига арильной группы в соседний сайт карбокатиона можно рассматривать как внутримолекулярное электрофильное замещение типа Фриделя-Крафтса. Приближение арильного кольца со стороны, противоположной отходящему азоту иона диазония, генерирует промежуточное соединение иона фенония (показано в скобках выше), структура которого подобна иону бензолония. В этих двух примерах диастереомерные реагенты предпочтительно приводят к диастереомерным промежуточным соединениям, даже несмотря на то, что анизильная группа имеет гораздо большую способность к миграции, чем фенил.Донорство электронной пары гидроксильным заместителем затем действует, открывая трехчленное кольцо этих промежуточных соединений, давая кетоновые продукты.
4. Анхимерная помощь
При измерении скорости сольволиза алкилгалогенидов и сложных эфиров сульфоновой кислоты наблюдаются любопытные влияния соседних заместителей. Например, этилхлорид, неопентилхлорид (2,2-диметилпропилхлорид) и 2,2,2-трифенилэтилхлорид представляют собой 1º-алкилхлориды, которые гидролизуются во влажной муравьиной кислоте до смесей спиртов и олефинов (S N 1 & E1 механизмы).Скорости реакции для этилхлорида и неопентилхлорида почти идентичны, но трифенильное соединение реагирует в 60 000 раз быстрее. Уравнения для последних двух сольволизов показаны на следующей диаграмме. Очевидно, что в обоих случаях первоначально образованный 1º-карбокатион перегруппировался до образования продукта, как показано при нажатии на диаграмму. Однако повышенная скорость фенилзамещенного соединения вызывает недоумение, особенно с учетом большей электроотрицательности фенильных групп по сравнению с метилом (обратите внимание, что дифенилуксусная кислота более чем в девять раз кислотнее изомасляной кислоты).Чтобы объяснить неожиданную реакционную способность 2,2,2-трифенилэтилхлорида, предполагается, что пи-электроны подходящей фенильной группы способствуют ионизации 1º-хлорида за счет образования связи со стороны, противоположной связи C-Cl, как показано щелкнув по диаграмме второй раз. Это внутримолекулярное взаимодействие соответствует последнему примеру в предыдущем разделе и аналогично внутримолекулярной реакции S N 2. Образовавшийся ион фенония немедленно откроется для 3º-карбокатиона, в котором вспомогательная фенильная группа переместится в соседнее положение.Таким образом, соседнее ароматическое кольцо ускоряет определяющую скорость (эндотермическую) стадию ионизации, влияние, называемое анхимерной поддержкой (греч .: anchi = сосед).
Следующие ниже энергетические профили этих реакций иллюстрируют последовательность событий. Обе реакции начинаются с начальной стадии ионизации, определяющей скорость, переходное состояние которой окрашено в розовый цвет. Энергия активации для этого этапа больше для неопентилхлорида, потому что он приводит к дискретному 1º-карбокатиону.С другой стороны, ионизация трифенилэтилхлорида происходит при содействии соседней фенильной группы, и образующийся ион фенония немедленно открывается в очень стабильный дифенил-3º-карбокатион. Второй этап сольволиза неопентилхлорида — это быстрая перегруппировка 1º-карбокатиона в изомерный 3º-карбокатион. Переходное состояние для этой перестановки окрашено в зеленый цвет. В обоих случаях промежуточный 3º-карбокатион в конечном итоге диспропорционирует до смеси продуктов замещения и элиминирования.Существенное отличие состоит в том, что ионизационное переходное состояние для неопентилхлорида страдает всеми недостатками, связанными с образованием 1º-карбокатиона; тогда как переходное состояние для ионизации трифенилэтилхлорида имеет пониженную энергию из-за его феноний-подобного характера.
Анхимерная помощь не только проявляется в усилении ионизации, но и влияет на стереохимический исход реакций. Примером может служить ацетолиз диастереомерных производных 3-фенил-2-бутанола.Этот спирт имеет два хиральных центра и, следовательно, имеет четыре стереоизомера в виде двух пар энантиомеров. Диастереомерные конфигурации называются эритро и трео в соответствии с их корреляцией с тетрозами эритрозой и треозой. Как правило, эритроизомеры могут принимать затененную конформацию, в которой идентичные или похожие заместители в двух стереогенных сайтах затмевают друг друга. Треоизомеры не могут принимать такую конформацию. На следующей диаграмме тозилатное производное одного энантиомера каждого диастереомера изображено в виде проекции Фишера.Эти изомеры сольволизовали в горячем растворе уксусной кислоты, забуференном ацетатом натрия, и определяли конфигурации образующихся эфиров ацетата. Как и ожидалось от процесса S N 1, также было получено некоторое количество продукта удаления E1. Примечательно, что каждый диастереомер превращается в его эквивалентный диастереомерный ацетат (сохранение конфигурации). Кроме того, эритросоединение сохраняет свою энантиомерную чистоту; тогда как треотозилат дает рацемический ацетат и сам рацемизируется во время реакции.Если бы в этих реакциях образовывался промежуточный карбокатион, смеси эритро- и треоацетатов можно было бы ожидать от обоих тозилатов, но среди продуктов были обнаружены только следовые количества противоположного диастереомера.
При нажатии на диаграмму будет продемонстрировано контролирующее влияние анхимерной помощи фенильной группы. Во-первых, молекула принимает конформацию, в которой фенильный заместитель ориентирован против тозилатной группы. Затем пара пи-электронов от бензольного кольца связывается с С2, когда тозилат-анион уходит, образуя промежуточное соединение фенония (в скобках).Промежуточный продукт из эритротозилата хиральный, а из треотозилата — ахиральный (обратите внимание на плоскость симметрии, разделяющую трехчленное кольцо пополам). В каждом случае C2 и C3 конституционально эквивалентны, и нуклеофильная атака ацетат-анионом происходит одинаково хорошо в любом положении (зеленые и голубые стрелки). В результате равных скоростей образования продукта за счет связывания ацетата с C2 и C3 ахиральный трео-промежуточный продукт дает 50:50 (рацемическую) смесь энантиомеров трео: (2R, 3S) из синих стрелок и (2S, 3R) из зеленые стрелки.Напротив, связывание ацетата с C2 и C3 промежуточного эритропродукта дает такой же энантиомер эритропродукта (2S, 3S). Поскольку первоначальная ионизация до фенониевых промежуточных продуктов обратима, мы не удивлены, обнаружив, что непрореагировавший эритротозилат не изменился; тогда как непрореагировавший треотозилат рацемизируется.
5. Анхимерная помощь со стороны других соседних групп
Способность пи-электронов в подходящем соседнем бензольном кольце способствовать ионизации C-X, где X представляет собой галоген или сложный сульфонатный эфир, была описана в предыдущем разделе.Другие ароматические кольца, такие как нафталин, фуран и тиофен, могут действовать аналогичным образом, как и пи-электроны двойных и тройных связей. На следующей диаграмме показаны три примера взаимодействия соседних двойных связей, первый из которых является одним из самых ярких случаев анхимерной помощи за всю историю наблюдений. Пунктирные линии показывают делокализацию заряда — обычная практика. Текстовое поле под диаграммой содержит дополнительные комментарии к этим примерам.
Примеры других возмущений соседних групп, включая помощь несвязывающей электронной пары соседними атомами серы, кислорода и азота, будут показаны выше при нажатии соответствующей кнопки под диаграммой.Комментарий текстового поля изменится в соответствии с примерами. В большинстве случаев с участием гетероатома образуется «ониевый» интермедиат, в котором гетероатом заряжен. Соседние атомы галогена могут также стабилизировать карбокатионы, как отмечалось ранее в отношении транс-анти добавок к циклическим алкенам. Сообщалось также о функциональной перестройке с помощью промежуточных соединений галония. Например, раствор диаксиального 2-бром-3-хлоростероида в хлороформе, показанный слева внизу, самопроизвольно перестраивается в более стабильный диэкваториальный 2-хлор-3-бром-изомер, изображенный справа.Перегруппировка обратима и происходит по циклическому иону бромония, указанному в скобках.
6. Неклассическая гипотеза карбокации
Роль промежуточных продуктов карбокатиона во многих органических реакциях хорошо известна. Некоторые, например трет-бутил, локализованы. Некоторые из них, такие как аллил и бензил, стабилизируются сопряжением с пи-электронными системами. Некоторые из них, как описано выше, стабилизируются за счет связывания с соседними нуклеофилами.Во всех случаях анхимерной помощи, описанной выше, частицы с делокализацией или перераспределением заряда являются промежуточными на пути реакции. Такие промежуточные продукты могут быть изолированы в некоторых случаях, но обычно они существуют только временно. Ускорение скорости ионизации объясняется структурным и энергетическим сходством переходных состояний с промежуточными продуктами, которые они производят (постулат Хаммонда).
Анхимерная помощь обычно связана с одной или несколькими из следующих наблюдаемых характеристик.
• Оцените ускорение по сравнению с аналогичными реакциями без помощи.
• Стереоэлектронный контроль, который приводит к различиям в скорости и продукте между стереоизомерами.
• Сохранение конфигурации в заменяемых продуктах.
• Рацемизация продуктов (и часто реагентов), когда участвует симметричный мостиковый промежуточный продукт.
Сольволиз экзо- и эндо-2-норборнилсульфонатных эфиров выявил различия, которые предполагают анхимерную помощь экзо-изомеру.Как показано на следующей диаграмме, скорость ацетолиза экзо-изомера значительно выше, чем скорость эндо-изомера, который реагирует со скоростью, аналогичной циклогексильному производному. Первая замена протекает с полным сохранением конфигурации и рацемизации; тогда как эндо-изомер замещен с инверсией конфигурации и сохраняет небольшую степень оптической активности. Источником этой помощи была предложена электронная пара сигма-связи C1: C6, которая идеально ориентирована против уходящей сульфонатной группы.Сигма-делокализованный ион (показан в скобках) был предложен в качестве промежуточного звена, отображаемого при нажатии на диаграмму. Поскольку этот мостиковый ион является симметричным, ожидается образование рацемического ацетата. Термин «неклассический» был применен к этому катиону с делокализацией заряда, поскольку он оказался уникальным.
Для сравнения, эндо-изомер ионизируется до классического 2º-карбокатиона, который быстро превращается в более стабильный неклассический ион. Некоторое количество ацетат-аниона может связываться с 2º-карбокатионом до того, как он изменится, что объясняет остаточную оптическую активность в этой реакции.
Не всех убедила такая интерпретация свидетельств. Главными сторонниками неклассической точки зрения были С. Винштейн и Дж. Д. Робертс. Основная оппозиция исходила от Г. К. Брауна, который придерживался более традиционной рационализации. Браун указал, что норборнильные соединения лучше по сравнению с циклопентилом, чем с аналогами циклогексила (затмевающий штамм), и в таком сравнении эндо-изомер аномально медленный, а экзо-изомер только в 14 раз быстрее, чем циклопентил.Рацемический продукт был объяснен предположением, что взаимное превращение энантиомерных классических карбокатионов происходит очень быстро по шкале времени реакции. Браун также отметил, что присоединение стабилизирующего арильного заместителя к C2 не снижает скорость увеличения экзоионизации или предпочтение образования экзо-продукта. Поскольку эти последние сольволизы протекают посредством бензильного катиона, предполагается, что содействие сигма-связи будет минимальным. Следовательно, повышение скорости и сохранение конфигурации становятся менее значимыми как неклассические индикаторы.Этот последний эксперимент, в котором арильным заместителем был п-анизил (An), изображен в левой части диаграммы ниже.
Несмотря на разрушительные аргументы Брауна, другие эксперименты предоставили дополнительную поддержку неклассической точке зрения. Как показано в правой части диаграммы, электроноакцепторные заместители на C6 ( 2 R) замедляли экзореактивность сильнее, чем эндореактивность. Аналогичный эффект отмечен для таких заместителей при C1 ( 1 R).Это влияние лучше всего объясняется неклассической гипотезой, согласно которой частичный положительный заряд должен нести C1, C2 и C6.
Интерпретации значительного количества доказательств, собранных на этом этапе, можно обобщить на диаграмме справа. На первом дисплее неклассический мостиковый катион показан как переходное состояние для взаимного превращения классических карбокатионов. Такое соотношение соответствует перегруппировке неопентилхлорида.Вторая возможность, представленная щелчком по диаграмме, имеет неклассический ион в качестве промежуточного звена с более высокой энергией, связывая классические ионы. Наконец, если щелкнуть диаграмму второй раз, можно увидеть возможность того, что неклассический ион представляет собой более стабильное промежуточное соединение.
К середине 1960-х химические методы и методы ЯМР были усовершенствованы до стадии, которая позволила прямое наблюдение карбокатионов в слабонуклеофильных кислых растворах, часто называемых «суперкислотами». Большая часть этой работы была проведена Джорджем Олахом (Нобелевская премия, 1994) с использованием смешанных растворителей, состоящих из SbF 5 , SO 2 , SO 2 F 2 и SO 2 FCl.При низких температурах были получены и интерпретированы спектры ЯМР 1 H и 13 C для (CH 3 ) 3 C (+) и (CH 3 ) 2 CH (+). Как и ожидалось, заряженный трехкоординатный атом углерода показал сигнал 13 C более 300 ppm в слабом поле от TMS. Когда аналогичные измерения ЯМР были применены к 2-норборнильному катиону, был обнаружен ряд быстрых протонных сдвигов. Их можно «заморозить», работая при низкой температуре, 3,2-смену при -70 ° C и более быструю 6,2-смену при -130 ° C.Результирующий спектр, который оставался неизменным при температурах до -160 ° C, не имел сигналов слабого поля, близких к ожидаемым для классического 2 °-карбокатиона, и свидетельствовал о неклассической структуре. Недавно твердотельные спектры ЯМР 13 C при 5 ° K подтвердили соответствие с неклассическим ионом. На основании этих и других спектроскопических исследований неклассический катион с сигма-мостиковым мостиком был твердо идентифицирован как более стабильная разновидность карбокатиона, имеющая 2-норборнильную структуру. Дальнейшее подтверждение было получено в 2013 году исследователями из Германии, проведенными с помощью тщательных рентгеновских кристаллографических измерений отожженной [C 7 H 11 ] + [Al 2 Br 7 ] — соли при 40º. К.
Существуют ли другие относительно стабильные неклассические карбокатионы? Было идентифицировано несколько, которые, кажется, соответствуют этой классификации, но немногие из них были изучены столь же исчерпывающе, как 2-норборнил. Один из лучших критериев для оценки ионов-кандидатов — установить, являются ли один или несколько участвующих атомов углерода гипервалентными (имеют более четырех координирующих групп). На следующей диаграмме простейший гипервалентный карбокатион, метаноний, изображен слева в серой заштрихованной рамке.Этот ион обычно наблюдается в масс-спектре метана (газовая фаза), но разлагается в растворе из-за его чрезвычайной кислотности. Справа находятся два более крупных неклассических иона, 2-норборнил и 7-норборненил. В каждом случае идентифицируется пятикоординированный атом углерода. Вкладчики резонанса в эти ионы показаны справа от пунктирной связи, и на всех рисунках делокализованная электронная пара окрашена в синий цвет. Наконец, широкий обзор этой классификации, предложенный Олахом в его Нобелевской лекции, будет показан при нажатии на диаграмму.
Посмотреть модель 2-норборнильного катиона.
Перегруппировки в электронодефицитные гетероатомы |
|---|
Перегруппировки в электронодефицитные гетероатомы
1. Перегруппировки катионного кислорода
Протонирование двухвалентного атома кислорода спиртов и простых эфиров сильными кислотами дает трехкоординатный оксониевый катион . Поскольку кислород иона оксония имеет октет валентной оболочки, он не является электронно-дефицитным сайтом и не может служить концом перегруппировки.Чтобы вызвать перегруппировку таким же образом, как трехкоординатный карбокатион, кислород должен быть преобразован в однокординатный оксакатион , как показано на следующей диаграмме. Сдвиг 1,2-алкила или арила затем превращает относительно нестабильный оксакатион в более стабильный карбокатион.
Простейшим предшественником окса-катиона является пероксид или эквивалентное производное (например, R-O-OH или R-O-X). Удаление гидроксид-аниона из гидропероксида является энергетически невыгодным, если он изначально не превращается в более удаляемую группу способом, аналогичным тому, который используется для облегчения реакций замещения спиртов.При протонировании гидроксильной группы уходящая группа становится водой, вызывая, таким образом, оксакатион. На этой стратегии основана полезная промышленная методика получения фенола (и ацетона).
Перегруппировка Байера-Виллигера
Катализируемая кислотой реакция кетонов с гидропероксидными производными известна как реакция Байера-Виллигера . Общее уравнение, иллюстрирующее эту реакцию окисления, показано ниже, и можно отметить, что стадия перегруппировки аналогична стадии перегруппировки пинакола.Сложные эфиры или лактоны являются основными продуктами кетоновых реагентов. В этом уравнении дискретный оксакатион изображен в качестве промежуточного звена, но более вероятно, что перегруппировка согласована, как будет показано при нажатии на уравнение. После добавления перкислоты к карбонильной группе перегруппировка может быть облегчена за счет внутримолекулярной водородной связи, как показано в скобках справа.
Способность к миграции различных групп заместителей (например, 1 R и 2 R) обычно составляет: 3º-алкил> 2º-алкил ~ бензил ~ фенил> 1º-алкил> метил .Стереоэлектронные факторы способствуют антиперипланарной ориентации мигрирующей группы по отношению к уходящей части и в некоторых случаях будут контролировать перестройку. Пример будет показан ниже при повторном щелчке по дисплею. Обмен надкислот с перуксусной кислотой приводит к внутримолекулярной реакции Байера-Виллигера посредством бициклического ацилпероксида, указанного в скобках. Здесь стереоэлектроника способствует миграции менее замещенного α-углерода. Лактонный продукт идентифицировали этерификацией и эфирным обменом с метанолом, получая метил-2-карбометокси-7-гидроксигептаноат.
Альдегиды обычно окисляются до карбоновых кислот в условиях, используемых для реакции Байера-Виллигера.
Хотя перекись водорода сама по себе может использоваться в реакции Байера-Виллигера, она может присоединяться на обоих концах к реакционноспособным карбонильным группам, давая циклические димерные, тримерные и соединения с более высоким присоединением. Следовательно, производные, такие как перкислоты (Z = RCO и ArCO выше), являются предпочтительными реагентами для этой реакции. Среди наиболее распространенных надкислот, используемых в этом отношении: перуксусная кислота, пербензойная кислота и мета -хлорпербензойная кислота (MCPBA).Четыре примера этой окислительной перегруппировки приведены на следующей диаграмме. В большинстве этих примеров мигрирующая группа сохраняет свою конфигурацию в ходе перегруппировки, как и ожидалось для согласованного процесса. В примере № 3 интересно, что миграция 3º -алкильной группы моста является предпочтительной по сравнению с возможным фенильным сдвигом.
Некоторые окислительные перегруппировки Байера-Виллигера
2. Перегруппировки катионного или электронодефицитного азота
Поскольку многие простые соединения азота являются основаниями, при протонировании они образуют «ониевые» катионы.Два таких катиона показаны слева (в синей рамке) ниже. Поскольку катионы аммония и иминия имеют октет электрона валентной оболочки азота, такой атом азота не может служить местом для нуклеофильной связи или концом миграции. Чтобы катион азота инициировал перегруппировку, он должен иметь незаполненную валентную оболочку, и два таких азакатиона показаны в центре диаграммы (розовая рамка). Электронно-дефицитный атом азота не обязательно должен быть катионом, как показывает пример нитрена справа (зеленая рамка).
Перегруппировка Бекмана
Если гидроксильная группа кетоксима удаляется под действием сильной кислоты или пентахлорида фосфора с последующим гидролизом, образуется амид. Полное удаление дериватизированной гидроксильной группы и ее пары связывающих электронов привело бы к двухвалентной sp-гибридизации азакатионов типа, изображенного на предыдущей диаграмме. Если бы это произошло, оба углеродных заместителя ( 1 R и 2 R) были бы кандидатами для последующего 1,2-сдвига.Однако на практике всегда группа, направленная против отходящего ОН, мигрирует в азот. Эта стереоспецифичность указывает на то, что 1,2-сдвиг согласован с расщеплением N-O, как показано ниже. Полученный промежуточный N-алкилированный нитрил будет реагировать с нуклеофилами (например, с водой) у электрофильного атома углерода, соседнего с «ониевым» азотом. Обратите внимание, что структура, нарисованная для этого промежуточного продукта, является более предпочтительной из двух вкладчиков резонанса, поскольку все тяжелые атомы имеют заполненные октеты валентной оболочки.Присоединение нуклеофила к атому азота потребовало бы расширения его валентной оболочки до десяти электронов или образования нестабильных диполярных частиц. Исходным продуктом гидратации на углероде является иминол, который немедленно таутомеризуется с образованием более стабильного амида. Реакции с PCl 5 , вероятно, дают иминохлорид, который, в свою очередь, гидролизуется до того же амида.
Первым примером в следующей группе реакций является типичная перегруппировка Бекмана.Оксим из циклогексанона имеет идентичные карбонильные заместители. и поэтому существует как единственный изомер. Продукт перегруппировки представляет собой лактам (циклический амид), который может быть гидролизован до омега-аминокислоты. Этот лактам служит важным промышленным предшественником нейлона 6. Второй пример включает производное оксима с различными карбонильными заместителями, которое существует в виде пары стереоизомеров (син и анти). Анти-изомер перестраивается за счет 1,2-фенильного сдвига, тогда как син-изомер претерпевает 1,2-изопропиловый сдвиг.Первая реакция протекает намного быстрее, чем вторая, предположительно потому, что она протекает через относительно стабильный промежуточный ион фенония (структура в заштрихованной рамке). Обратите внимание, что уходящая пикратная группа (2,4,6-тринитрофенолят) является стабильным анионом. Пример № 3 — еще один случай, демонстрирующий стереоспецифичность перегруппировки Бекмана. 1,2-сдвиг ортофенольного заместителя происходит быстрее, чем сдвиг незамещенной фенильной группы, и гидроксил идеально расположен для связи с электрофильным углеродом промежуточного соединения.Следовательно, продукт анти-изомера представляет собой бензоксазольный гетероцикл.
Четвертый пример, приведенный выше, показывает необычное расхождение в поведении, которое иногда происходит, когда мигрирующие фрагменты заместителя от промежуточного соединения оставляют нитрил в качестве стабильного продукта. Это было названо ненормальной реакцией Бекмана.
Жесткая конфигурация катиона фенония, показанная выше, налагает структурные ограничения, которые хорошо демонстрируются скоростями перегруппировки некоторых бициклических соединений с конденсированным кольцом.Щелкнув по диаграмме, вы увидите результаты такого исследования. Исследованы оксимные производные фенилкетонов, входящие в шестичленные, семичленные и восьмичленные конденсированные кольца. Из-за углеродной цепи, соединяющей оксимную функцию с орто-углеродом бензольного кольца, ион фенония, который обычно способствует миграции фенила, может не иметь своей предпочтительной структуры (трехчленное кольцо, ортогональное фенильному кольцу). Трехуглеродная мостиковая цепь для n = 6 является таким случаем, и перегруппировка анти-изомера происходит очень медленно.По мере увеличения длины мостиковой цепи ее ограничение становится менее жестким, и скорость перегруппировки увеличивается. Восьмичленный пикрат оксима быстро гидролизуется с образованием девятичленного лактама с высоким выходом.
R 2 C = N-NH 2 + HNO 2 RCONHR + N 2 |
Перегруппировки типа Beckmann также могут быть выполнены путем обработки гидразонов азотистой кислотой. , как показано справа.Как правило, это менее желательная процедура, поскольку чистые гидразоны получить труднее, а выход чистого продукта ниже.
Прямая перегруппировка кетонов, позволяющая избежать необходимости получения производных, возможна с помощью процедуры, известной как перегруппировка Шмидта . Катализируемое кислотой присоединение азотной кислоты к карбонильной группе кетона создает нестабильный азидокарбинол, который при дегидратации дает тот же триазониевый катион, предположительно образующийся в качестве промежуточного соединения при дезаминировании азотистой кислоты гидразона.Перегруппировка этого вида путем быстрой потери азота затем инициирует перегруппировку, подобную Бекману. Если щелкнуть верхнюю диаграмму второй раз, будут представлены два примера перегруппировки Шмидта.
Перегруппировка Стиглица
Примеры перегруппировок в sp 2 гибридизированных азакатионов относительно редки по сравнению с их углеродными аналогами. Исходными материалами для получения таких азакатионов обычно являются хлорамины или гидроксиламины. Ниже показаны четыре примера этих преобразований, иногда называемых «перестановками Штиглица».Первый пример похож на перегруппировку Вагнера-Меервейна, а второй — на пинаколовую перегруппировку. Конкурентные сдвиги пара-замещенных фенильных групп в реакциях, подобных примеру № 3, демонстрируют, что метокси-заместитель облегчает перегруппировку, тогда как нитрозаместитель ее замедляет. В последнем примере сопряженный азакатион активирует бензольное кольцо до нуклеофильного замещения, в отличие от обычной роли аминовых заместителей.
Перегруппировка ацилнитренов в изоцианаты
Несколько полезных и общих процедур разложения производных карбоновой кислоты до аминов включают перегруппировку ацилнитрена в изоцианат.Хотя атом азота нитрена не имеет формального заряда, он электронно-дефицитный и служит локусом для 1,2-перегруппировок. Как показано на следующей диаграмме, ацилнитрены могут быть получены из различных амидоподобных исходных соединений. После образования ацилнитрены быстро перегруппировываются в относительно стабильные изомеры изоцианата, которые можно выделять или подвергать взаимодействию с гидроксильными растворителями. Наиболее частое применение этой перегруппировки — синтез аминов. Таким образом, добавление воды к кетеноподобным изоцианатам дает нестабильную карбаминовую кислоту, которая разлагается на амин и диоксид углерода.Общие процедуры получения предшественников нитрена перечислены под диаграммой.
Hofmann Route: Первичные амиды превращаются в N-галогенированные производные под действием HOX или X 2 в щелочном растворе. Избыточное основание образует сопряженное основание продукта.
Lossen Route: Производное гидроксамовой кислоты (RCONHOH) получают реакцией сложного эфира с гидроксиламином. Гидроксамовая кислота подвергается O-ацилированию, а затем превращается в ее конъюгированное основание.
Курциус Маршрут: Ацилазид (RCON 3 ) получают одним из двух способов. (i) Реакция ацилхлорида с азидом натрия или (ii) Реакция сложного эфира с избытком гидразина с последующей реакцией ацилгидразидного продукта (RCONHNH 2 ) с холодной азотистой кислотой. Ацилазиды при нагревании разлагаются до изоцианатов.
Маршрут Шмидта: Вариант процедуры Курциуса, в котором карбоновая кислота нагревается с азойной кислотой (HN 3 ) и кислотным катализатором.
Некоторые примеры этих различных перестановок показаны на следующей диаграмме. В каждом случае зеленая заглавная буква (C, H, L или S) обозначает тип реакции. Первые три реакции иллюстрируют перегруппировку Хофмана, которая является особенно полезным методом получения аминов. В последнем примере показана реакция Курциуса на сложный диэфир в виде промежуточного бис-ацилгидразида. Также показан альтернативный подход Куртиуса к аминовому продукту из примера №2.Процедура Куртиуса особенно полезна, если желаемым продуктом является изоцианат, поскольку разложение промежуточного ацилазида можно проводить в отсутствие гидроксильных растворителей, которые могли бы реагировать с изоцианатом.
Еще пять примеров будут отображены выше, если щелкнуть диаграмму. Пример № 5 показывает реакцию Шмидта, в которой оптически активная карбоновая кислота является субстратом. S-конфигурация мигрирующей фенэтильной группы сохраняется в аминовом продукте, подтверждая внутримолекулярный характер этих перегруппировок.Сохранение конфигурации наблюдается также в перегруппировках Курциуса, Лоссена и Гофмана. Реакции №6 и №7 представляют собой интересные случаи, в которых вода отсутствует во время образования и реакции изоцианата. Спиртовой растворитель в №6 добавляется к изоцианату с образованием сложного эфира карбамата, известного как уретан . В отличие от нестабильных карбаминовых кислот уретаны не разлагаются и могут быть выделены в виде чистых соединений. Если бы вода была растворителем, полученный 1º-енамин перегруппировался бы в имин и гидролизовал бы до альдегида.В перегруппировке Лоссена (№7) бутиламин добавляется к изоцианату с образованием замещенной мочевины (мочевина — NH 2 CONH 2 ).
Перегруппировка Хофмана в реакции № 8 представляет собой новый пример таутомерии ацетиленового 1º-амина в нитрил. Наконец, последний пример иллюстрирует селективную перегруппировку Гофмана бромимида. Реакционная способность пара карбонильной группы к электроноакцепторному нитрозаместителю увеличивается по сравнению с другим имид-карбонилом.Следовательно, здесь предпочтительно происходит катализируемый основанием гидролиз, оставляя ацилнитреновый фрагмент мета для нитрофункции.
Перегруппировки ацилкарбенов |
|---|
Перегруппировки ацилкарбенов
1. Реакция Арндта-Эйстерта
Перегруппировка ацилнитренов в изоцианаты, которая является сутью перегруппировок Хофмана, Курциуса и Лоссена, проходит параллельно с перегруппировкой ацилкарбенов в кетены, превращение, называемое перегруппировкой Вольфа .Эта перегруппировка является критическим этапом в процедуре Arndt-Eistert для удлинения карбоновой кислоты одним метиленовым звеном, как описано на диаграмме ниже. Исходная кислота, указанная слева, превращается сначала в производное ацилхлорида, а затем в диазометилкетон. Диазометан имеет нуклеофильную метиленовую группу, как показано резонансными формулами, нарисованными в заштрихованной рамке. Ацилирование метиленового углерода дает равновесную смесь разновидностей диазония и диазометилкетона плюс хлористый водород (указано в скобках).Если HCl не нейтрализуется основанием, эта смесь реагирует дальше с образованием хлорметилкетона с потерей азота. Однако, если HCl нейтрализуется по мере его образования, получается относительно стабильный диазокетон, который может быть использован в последующих реакциях.
Поскольку сам диазометан может действовать как основание, ход данной реакции определяется способом, которым соединяются реагенты. Когда эфирный раствор диазометана медленно добавляют к теплому раствору хлорангидрида, наблюдается выделение азота, и хлорметилкетон является основным продуктом.Для этой реакции требуется один эквивалент диазометана. Если добавить обратное, так что холодный раствор хлорангидрида медленно добавляют к избытку диазометана в холодном растворе эфира, снова наблюдается выделение азота; но расходуются два эквивалента диазометана. Продуктами реакции диазометана с HCl являются диазокетон и метилхлорид (газ).
Для проведения реакции Арндта-Эйстерта диазокетон разлагают в присутствии серебряного катализатора (обычно AgO 2 или AgNO 3 ) вместе с теплотой или световой энергией.Результирующая перегруппировка Вольфа генерирует кетен, который быстро реагирует с любыми гидроксильными или аминными реагентами, которые могут присутствовать в растворе. Общие уравнения слева внизу показывают, что конечный продукт реакции Арндта-Эйстерта может быть карбоновой кислотой, сложным эфиром или амидом.
Три конкретных примера этой процедуры представлены справа от общих уравнений. Первые два примера представляют собой типичные реакции Арндта-Эйстерта. Переносимость других функциональных групп, таких как нитро, демонстрируется первым случаем; и второй пример показывает, что конфигурация мигрирующей группы сохраняется при перегруппировке.Другие реакции кетена, такие как [2 + 2] циклоприсоединение, могут иметь место в отсутствие гидроксильных растворителей или аминов. Реакция №3 является примером такой альтернативной реакции. Новые связи в цикладдукте окрашены в розовый цвет.
2. Реакции диазокетонов
Реакция Арндта-Эйстерта является частным случаем более общего класса реакций диазокетона. Если предположить, что диазокетоны обычно разлагаются до ацилкарбенов, то можно представить себе множество последующих реакций, и многие из них были реализованы.На следующей диаграмме показаны некоторые из этих превращений, происходящие из формулы диазокетона в центре диаграммы. Молекулярный азот теряется в каждом случае, и путь перегруппировки Вольфа находится слева. Большинство других реакций отражают способность карбенов вставлять сигма-связи или присоединяться к двойным связям. Металлические катализаторы иногда используются для облегчения определенных реакций и могут связываться с карбенами с образованием карбеноидных промежуточных продуктов.
При нажатии на диаграмму будут показаны некоторые примеры этих реакций диазокетона.
Практические задачи |
|---|
Здесь представлены некоторые проблемы, связанные с молекулярными перегруппировками. |
Эта страница является собственностью Уильяма Ройша.
Комментарии, вопросы и ошибки следует
направлять по адресу [email protected].
Эти страницы предоставляются IOCD для оказания помощи в наращивании потенциала в области химического образования. 05.05.2013
Перестройка гена TEL при остром лимфобластном лейкозе: новый генетический маркер с прогностическим значением
Цель: Перестройки гена TEL из-за транслокации хромосом 12; 21 являются наиболее частой молекулярно-генетической аномалией при остром лимфобластном лейкозе (ОЛЛ) у детей, встречающейся примерно в 25% случаев с иммунофенотипом B-предшественника.Ограниченное количество клинически полезных генетических маркеров этого подтипа лейкемии побудило нас оценить статус TEL как предиктор результата лечения.
Пациенты и методы: Мы определили статус гена TEL (реаранжированный или зародышевый) в 188 случаях острого лейкоза-предшественника B с помощью анализа саузерн-блоттинга и связали результаты с выживаемостью без событий. Все сравнения результатов были стратифицированы по схеме лечения, классификации риска, возрасту и количеству лейкоцитов.
Полученные результаты: У 48 пациентов (26%) был реаранжированный ген TEL. Через 5 лет наблюдения примерно 91% +/- 5% (SE) этой группы были выжившими без событий, по сравнению только с 65% +/- 5% группы с TEL зародышевой линии (стратифицированный логарифмический ранг P = 0,011). Для негипердиплоидных пациентов отношение шансов неблагоприятного события в группе TEL зародышевой линии к таковому для группы реорганизованной TEL составляло 4.06 (95% доверительный интервал от 1,86 до 8,84). Связь перестройки TEL с благоприятным прогнозом не зависела от признанных признаков хорошего риска при лейкемии-предшественнике B, включая возраст, исходное количество лейкоцитов и гипердиплоидию.
Заключение: Перестройка гена TEL отличает большую группу детей с благоприятным прогнозом лейкоза B-предшественника, которые не могут быть идентифицированы стандартными прогностическими признаками.Этих пациентов можно лечить менее агрессивно без потери терапевтической эффективности.
Соматическая перестройка в B-клетках: это (в основном) Nuclear Physics
Cell. Авторская рукопись; доступно в PMC 2016 13 августа.
Опубликован в окончательной отредактированной форме как:
PMCID: PMC4537798
NIHMSID: NIHMS710323
Эрез Либерман Эйден
1 Центр архитектуры генома, Бэй , TX77030, USA
Rafael Casellas
2 Genomics & Immunity, NIAMS and NCI, National Institutes of Health, 10 Center Drive, Bethesda, MD 20892, USA
1 Центр архитектуры генома, Колледж Бейлора Medicine, Houston, TX77030, USA
2 Genomics & Immunity, NIAMS и NCI, National Institutes of Health, 10 Center Drive, Bethesda, MD 20892, USA
Окончательная отредактированная версия этой статьи доступна на сайте CellSee, другие статьи в ЧВК, цитирующих опубликованную статью.Abstract
Мы обсуждаем, как принципы ядерной архитектуры управляют типичными перестройками генов в B-лимфоцитах, тогда как горячие точки транслокации и рецидивирующие поражения отражают степень AID-опосредованного повреждения и отбора ДНК.
Хромосомные перестройки необходимы для развития человека. В локусах гена иммуноглобулина ( Ig ) в B-клетках костного мозга белки гена активации рекомбинации (RAG) опосредуют соединение сегментов V, D и J для создания разнообразного набора антител.На периферии индуцированная активацией цитидиндезаминаза AID инициирует рекомбинацию переключения классов (CSR) константных доменов Igh , приводя к различным изотипам антител. Эти строго регулируемые процессы лежат в основе адаптивного иммунного ответа.
Ненаправленные перестройки также важны для клеточного гомеостаза и целостности генома. Спонтанные двухцепочечные разрывы ДНК (DSB) обычно репарируются в цис посредством негомологичного соединения концов (NHEJ) или в транс посредством гомологичной рекомбинации (HR).Хотя эти процессы в значительной степени полезны, эти процессы иногда генерируют вызывающие рак транслокации, которые сопоставляют онкогены (например, myc ) с мощными энхансерами Ig .
Мы рассматриваем три принципа ядерной архитектуры, которые влияют на паттерны рекомбинации в В-клетках: сворачивание полимера, образование петель между конвергентными мотивами CTCF и компартментализация A – B. В то время как рекомбинация гена Ig развивалась для использования ядерной архитектуры, повреждение ДНК и отбор вызывают повторяющиеся патологические перестройки в онкогенезе В-клеток.
Сворачивание полимеров хроматина и его роль в CSR
Простая оценка частоты перестройки между двумя локусами, A и B, представляет собой вероятность того, что они находятся в непосредственной пространственной близости внутри ядра клетки. В отсутствие локальных особенностей сворачивания (таких как петли) эта вероятность должна монотонно снижаться, поскольку A и B расположены дальше друг от друга по контуру генома. Такое снижение легко увидеть с помощью экспериментов Hi-C (Lieberman-Aiden et al., 2009). Снижение проходит через несколько режимов масштабирования. Для расстояний между локусами от 500kb до 7Mb ранние эксперименты Hi-C показали, что частота контакта связана с расстоянием по степенному закону с показателем степени примерно -1,0.
Теория полимеров дает обоснование этим наблюдениям. В таких исследованиях теоретическое и физическое моделирование конденсированного хроматина используется для вывода взаимосвязи между одномерной последовательностью и трехмерной близостью в пространстве. В настоящее время наиболее часто используемой моделью является фрактальная глобула, которая предсказывает, что частота контакта (или рекомбинации) между двумя локусами примерно пропорциональна расстоянию между ними (Lieberman-Aiden et al., 2009). Полученные прогнозы близко соответствуют эмпирическим значениям, полученным Hi-C.
Многие исследования подтвердили силу полимерных моделей в прогнозировании профилей рекомбинации. Простейшее предсказание, что большинство DSB должно быть разрешено в cis , было подтверждено исследованиями, в которых разрывы были вызваны RAG или ионизирующим излучением. Более количественные прогнозы были сделаны с использованием секвенирования следующего поколения для картирования перестроек. Эти методы основаны на встраивании сайтов рестрикции для I-SceI, эндонуклеазы хоминга дрожжей с редким мотивом длиной 18 п.н., в представляющие интерес локусы, такие как Igh и myc .Путем трансдукции активированных В-клеток с помощью I-SceI, DSBs формируются в целевых локусах, и результирующие перестройки могут быть профилированы высокопроизводительным способом (Chiarle et al., 2011; Klein et al., 2011). Монотонное снижение частоты рекомбинации I-SceI характеризовалось масштабированием по степенному закону (-1,3), аналогичным оценкам, полученным с помощью Hi-C и полимерного моделирования. Таким образом, геномы млекопитающих перестраиваются в cis с профилем, который имитирует полимерное поведение хроматина. (Обратите внимание, что недавние улучшения в картах Hi-C, вероятно, приведут к значительным улучшениям в лежащих в основе полимерных моделях.)
CSR явно выигрывает от этой склонности. Во время CSR активированные B-клетки заменяют константный домен IgM (Cμ) доменом нижележащего изотипа (Cγ, Cα или Cε). У мыши C H ограничены относительно небольшой областью (160 КБ) внутри обширного локуса Igh (2,8 МБ,). Рекомбинации способствует транскрипция областей переключателя (S) перед каждым доменом C H , что придает доступность AID и приводит к DSB. В экспериментах, в которых области переключения были заменены сайтами I-SceI (Gostissa et al., 2014; Zarrin et al., 2007), индукция разрывов I-SceI способствовала CSR. Проксимальные разрывы I-SceI рекомбинированы на более высоких частотах, чем дальние, что согласуется с идеей о том, что, по крайней мере частично, монотонное снижение взаимодействий Igh влияет на эти перестройки. Таким образом, CSR, по-видимому, использует полимерное поведение хромосом, вызывая события рекомбинации между областями переключения посредством репарации DSB, которые приходят в пространственную близость.
Паттерны перестройки часто отражают принципы ядерной архитектуры(A) Распределение мотивов связывания CTCF (красные и синие полосы) в локусе Igh .Положение каждого сайта определяется из списка пиков ChIP-Seq (Choi et al., 2013). Необработанный сигнал был слабым на нескольких аннотированных пиках, которые поэтому были удалены вручную. Стрелки и цвета обозначают ориентацию мотивов CTCF (вперед = красный, обратный = синий). Положение сайтов CTCF и сегментов V, D и J в локусе отрисовано в масштабе. Поразительно, что V-область содержит 109 мотивов CTCF, все в прямой ориентации. Все они сталкиваются с одним обратным мотивом около области D. На рисунке показан пример возможной петли между прямым мотивом CTCF и обратным мотивом.Обратите внимание, что, in vivo , такое образование петель можно было бы ожидать в присутствии реаранжированного DJ H , но локус показан в неупорядоченном состоянии, отражая состояние большинства клеток во время фактического эксперимента. Примечательно, что суперанкор, содержащий 10 мотивов CTCF в обратной ориентации, лежит в энхансере 3’RR; эти мотивы сталкиваются с одним прямым CTCF, также в локусе D. (B) Конфокальная микрофотография сравнения покоящихся В-клеток с активированными 72 часа. Образцы окрашивали анти-α-тубулином (красный) и DAPI (синий).После активации ядра становятся намного крупнее. (C) Корреляция между компартментализацией A – B (определяется собственным вектором Hi-C, синий), взаимодействием с Igh , измеренным с помощью 4C (красный), случайными хромосомными транслокациями с участием Igh и хромосомы 17 в отсутствие СПИДа (желтый) и при его наличии (зеленый). В клетках AID + / + наблюдается повторяющаяся транслокация горячих точек. Также показана карта Hi-C с разрешением 250 КБ. Данные взяты из (Hakim et al., 2012) и (Rao et al., 2014). (Г) Igh 4C-Seq профиль. Выделены семь (из 236) генов-мишеней AID, включая Il4ra и Gpr132 . Показано общее количество транслокаций Igh в каждом гене, как определено TC-Seq (Klein et al., 2011).
Дальнодействующее образование петель CTCF облегчает V (D) J-рекомбинацию и обеспечивает разнообразие антител.
V (D) J-рекомбинация происходит в более широком диапазоне расстояний, чем CSR. Первое событие рекомбинации объединяет сегменты D и J H , разделенные максимальным расстоянием 100 КБ (в геноме мыши).Напротив, рекомбинация V H -DJ H удаляет по крайней мере 45–150 КБ для наиболее проксимального сегмента V (IghV5-1) и до ~ 2,6 МБ для наиболее дистального (IghV1-86,). Таким образом, если рекомбинация V H -DJ H основывалась исключительно на монотонном поведении полимеров хроматина, проксимальные сегменты V H доминировали бы в репертуаре зрелых Ig, резко сокращая разнообразие антител. Однако микроскопические исследования показали, что весь локус Igh претерпевает конформационные изменения во время рекомбинации V (D) J (Jhunjhunwala et al., 2008). В частности, весь домен V H ассоциируется с сегментами D-J H в клетках, подвергающихся рекомбинации V H -DJ H (обзор (Subrahmanyam and Sen, 2012)). Ясно, что действуют механизмы, выходящие за рамки локального сворачивания полимера.
Недавняя Hi-C карта В-лимфобластоидных клеток человека пролила свет на эти механизмы. Исследование выявило ~ 10 000 петель между парами сайтов CTCF, чьи связывающие мотивы лежат в конвергентной ориентации (то есть обращены друг к другу, (Rao et al., 2014)). Локус Igh является яркой иллюстрацией этого принципа. Его область V H содержит> 100 сайтов CTCF, все из которых направлены вниз по течению. Они обращены к единственному сайту CTCF, направленному вверх по течению, который расположен на 5′-конце области D. (). Эта конфигурация согласуется с моделью, в которой зацикливание CTCF облегчает рекомбинацию между удаленными V H s и перестроенными DJ H s. (Подобные конфигурации наблюдаются в других локусах Ig ). В подтверждение этого, делеция сайтов связывания CTCF в области D уменьшала рекомбинацию с дистальными V H s (Guo et al., 2011; Lin et al., 2015). Таким образом, рекомбинация V (D) J может отличаться от CSR, поскольку она основана на CTCF-опосредованном образовании петель, а не только на сворачивании полимера. Помимо CTCF, другие факторы также участвуют в сокращении локуса Igh , включая E2A, Pax5, cohesin, Brg1 и YY-1 (rev. (Alt et al., 2013)). Как эти факторы дополняют CTCF-опосредованное образование петель, остается неясным.
Компартментализация A – B формирует случайные перестройки
DSB, которые образуются во время рекомбинации гена Ig , не всегда должным образом репарируются в цис и вместо этого могут приводить к случайным перестройкам как в цис , так и особенно в транс .В опухолях В-клеток от человека и других видов хромосомные транслокации часто соединяют онкогены с мощными энхансерами Ig , которые нарушают их экспрессию. Происхождение этих транслокаций обсуждается в течение нескольких десятилетий; близость, паттерны формирования DSB и выбор были предложены в качестве движущего механизма.
Сначала связь между архитектурой ядра и транслокациями была исследована с помощью микроскопии. Эти исследования проверяли гипотезу о том, что онкогены и локусы часто транслоцируются, потому что они преимущественно связываются в ядрах В-клеток.Флуоресцентная гибридизация in situ (FISH) показала, что Igh и myc находятся в пределах 1 мкм друг от друга во фракции активированных В-клеток, но частота перекрытия сильно варьировалась в разных исследованиях. В отчетах, где В-клетки анализировались вскоре после активации (<15 минут), Igh и myc , по-видимому, перекрывались более чем в 20% лимфоцитов (Osborne et al., 2007), тогда как на более поздних стадиях активации (72 часа) ) или в В-клетках зародышевого центра значения находились в диапазоне 3-5% (Gramlich et al., 2012; Wang et al., 2009). Эти эксперименты были интерпретированы как предположение, что, по крайней мере, при некоторых условиях Igh и myc предпочтительно связываются в В-клетках.
Однако в исследованиях не учитывалась частота случайного перекрытия локусов генов. Для 3D-FISH в диплоидном ядре с использованием порога перекрытия 1 мкм частота случайного перекрытия определяется как объем двух сфер радиусом 1 мкм (~ 8,4 мкм 3 ), деленный на общий объем ядра.Для В-клеток в состоянии покоя или вскоре после активации (15 минут) типичным является объем 55 мкм 3 (), что означает, что два гена случайным образом перекрываются в 15% клеток. Напротив, средний объем активированных В-клеток> 24 часа после активации составляет ~ 250 мкм 3 (), что приводит к случайной частоте перекрытия примерно 3,5%. (Для 2D FISH вероятность составляет 14% до активации и 5% после нее.) Эти соображения предполагают, что сообщаемые различия в перекрытии Igh-myc могут быть объяснены различиями в объеме ядра, а не предпочтительной ассоциацией.
Недавние эксперименты по картированию контактов дали более точные результаты. В частности, Hi-C-картирование с разрешением в килобаз не выявило пиков частоты контактов между парами локусов, лежащих в trans (Rao et al., 2014). Сходным образом, исследование 4C обнаружило, что Igh и myc не образуют предпочтительных локус-специфических ассоциаций в trans (Hakim et al., 2012). Важно отметить, что наблюдение, что нет никаких смещений в trans между отдельными локусами гена, не означает, что нет никаких смещений в trans вообще.Существование хромосомных территорий и преимущественная связь между ними хорошо известны; как и тот факт, что A – B-компартментализация хроматина приводит к пространственной сегрегации открытых и закрытых геномных доменов (Lieberman-Aiden et al., 2009; (de Laat and Grosveld, 2007). Следовательно, Igh и myc , расположены на хромосомах 12 и 15 соответственно, пространственно связаны с транскрипционно активными доменами по всему геному на сходных частотах (, (Hakim et al., 2012)).В соответствии с этой точкой зрения, высокопроизводительные FISH-эксперименты показывают, что частота перекрытия (<1 мкм) в В-клетках между Igh и типичным активным геном составляет 3-5%, тогда как частота перекрытия между Igh и любым молчаливым геном локус составляет 1-2% (Hakim et al., 2012).
Эти широкие паттерны пространственной близости влияют на паттерны перестройки. В AID — / — B-клетках или в облученных про-B-клетках — где распределение разрывов ДНК относительно однородно по геному — паттерны транслокации отражают как территории хромосом, так и компартментализацию A – B (Hakim et al., 2012). Предположительно, данные перегруппировки с более высоким разрешением будут отражать 6 субкомпартментов хроматина (A1 / A2 и B1 / B2 / B3 / B4), которые подразделяют исходные компартменты A и B и которые видны на картах Hi-C с самым высоким разрешением (Rao et al. ., 2014). Таким образом, в отсутствие повторяющихся повреждений ДНК случайная перестройка — те, которые не возникают из-за целевых механизмов, таких как рекомбинация VDJ, — в основном следуют широким контурам ядерной компартментализации.
Паттерны повреждений ДНК и выборочные паттерны формы давления при повторяющихся перестройках
Обсуждаемые до сих пор находки подчеркивают значительное влияние ядерной архитектуры на паттерны перегруппировки.Однако архитектура — не единственный фактор, влияющий на перестановку. Другой ключевой особенностью является частота ДНК DSB в определенных локусах. В присутствии ферментов, повреждающих ДНК, таких как RAG и AID, скорость образования таких повреждений не является постоянной по всему геному, а вместо этого нацелена на определенные участки. При раке необходимо учитывать еще один фактор: онкогенез может выбирать за или против определенных перестроек.
Горячие точки AID-опосредованного повреждения ДНК становятся горячими точками перестройки
Основная роль AID-опосредованного дезаминирования состоит в том, чтобы способствовать образованию DSB во время CSR.Тем не менее, AID также является важным фактором, способствующим перераспределению горячих точек. Эта тенденция четко наблюдалась в активированных В-клетках и В-клетках зародышевого центра. В дополнение к перестройкам, наблюдаемым на фоне AID — / — , активированные B-клетки с интактным AID обнаруживают горячие точки транслокации на myc и других онкогенах, участвующих в трансформации B-клеток. Путем мониторинга накопления факторов репарации RPA и Rad51 в резецированных разрывах ДНК было подтверждено, что эти горячие точки являются участками повторяющихся повреждений ДНК, вызванных AID (Hakim et al., 2012). В самом деле, существует прямая пропорциональность между степенью опосредованного AID образования DSB в горячих точках и абсолютным числом Igh транслокаций. Эти частоты перемещений значительно превышают прогнозы, основанные только на ядерной архитектуре. Например, Igh в 20 раз чаще пространственно совмещается с Gpr132 , который находится на расстоянии 400 килобайт на хромосоме 12, чем с Il4ra , который находится в trans на хромосоме 7 ().Но Igh в 20 раз чаще перестраивается с Il4ra , чем с Gpr132 . Точно так же myc вряд ли будет пространственно совмещаться с Igh : более 2000 генов с большей вероятностью будут это делать. Тем не менее, myc является одним из относительно небольшой фракции генов, которые периодически перестраиваются в Igh (Hakim et al., 2012).
Таким образом, паттерны AID-опосредованных DSBs являются основным фактором в определении сайтов рекуррентной транслокации в B-клетках, независимо от топологии.Это также относится к транслокациям, вызванным другими формами повторяющегося повреждения ДНК, такими как активность Rag1 / 2 или система CRISPR-Cas9 (Frock et al., 2015). Эти аргументы подчеркивают необходимость понимания факторов, которые делают локусы уязвимыми для каждого вида повреждения. В случае AID было сделано несколько предложений, включая наличие суперэнхансеров, транскрипцию конвергентных генов и высокую взаимосвязь между регуляторными элементами (обзор (Alinikula and Schatz, 2014)).
Архитектура ограничивает выбор вызывающих рак перестроек
В опухолях скорость, с которой обнаруживается конкретная перестройка, не отражает скорость ее образования в исходной клетке опухоли.Вместо этого существует сильный положительный отбор для случайных транслокаций, которые нарушают регуляцию экспрессии онкогенов. Например, транслокации Il4ra образуются в 10 раз чаще, чем транслокации myc в активированных В-клетках (), но — в отличие от myc — транслокации Il4ra в опухолях В-клеток еще не зарегистрированы.
При транслокациях Ig дерегуляция онкогенов имеет тенденцию быть результатом мощных энхансеров Igκ , Igλ или Igh .Поскольку эти энхансеры могут работать на расстоянии, считается, что они полагаются на пространственную близость, чтобы управлять своей биологической функцией. Однако эта же архитектура накладывает ограничения на их деятельность. Например, когда мышиные В-клетки без AID-опосредованного повреждения культивировали в неселективных условиях, случайные транслокации помещали 3′-энхансер Igh , известный как 3’RR, в широком диапазоне расстояний от myc (Ковальчук и др. др., 2012). После отбора (во время плазмоцитомагенеза мыши) тот же элемент был обнаружен не более чем на 500 т.п.н. из myc .За пределами этой точки не наблюдалось никаких локальных изменений в уровне рекрутирования PolII, экспрессии генов или в присутствии различных эпигенетических модификаций. Способность 3’RR активировать транслоцированные гены, которые лежат далеко по контуру генома, также наблюдалась в других зрелых B-клеточных опухолях (Gostissa et al., 2009). Учитывая вышеупомянутые данные об ориентации CTCF, вполне вероятно, что трансформационная способность энхансера 3′RR связана с тем фактом, что он расположен рядом с тем, что мы называем «суперанкор» CTCF: серией из 10 сайтов CTCF, все из которые указывают в том же направлении.Ключевое значение состоит в том, что функциональная досягаемость 3′RR и его суперанкоря — по крайней мере, в условиях транслокаций myc — составляет примерно половину мегабазы.
Таким образом, ядерная архитектура не только влияет на расположение и частоту перегруппировок, но также влияет на то, какие из этих событий выборочно благоприятствуют во время трансформации. Это наблюдение может иметь важные последствия для опухолей, которые не происходят из В- и Т-клеток и которые обычно не имеют целевого, рецидивирующего повреждения ДНК.
За последние 5 лет технический прогресс помог решить многие давние проблемы, связанные как с физиологической, так и с патологической перестройкой В-лимфоцитов. По мере развития области трехмерной геномики мы ожидаем, что она продолжит освещать эти механизмы и их значение для туморогенеза как внутри иммунной системы, так и за ее пределами.
Благодарности
Мы благодарим Воутера де Лаата, Неву Дюран, Сухаса Рао и Адриана Санборна за полезные обсуждения, а также Вольфганга Реша, Марей Дозе, Сухаса Рао и Сигрид Кнемайер за помощь в создании рисунка.
Сноски
Заявление издателя: Это PDF-файл неотредактированной рукописи, принятой к публикации. В качестве услуги для наших клиентов мы предоставляем эту раннюю версию рукописи. Рукопись будет подвергнута копирайтингу, верстке и рассмотрению полученного доказательства, прежде чем она будет опубликована в окончательной форме для цитирования. Обратите внимание, что во время производственного процесса могут быть обнаружены ошибки, которые могут повлиять на содержание, и все юридические оговорки, относящиеся к журналу, имеют отношение.
Ссылки
- Alinikula J, Schatz DG. Клетка. 2014; 159: 1490–1492. [PubMed] [Google Scholar]
- Alt FW, Zhang Y, Meng FL, Guo C, Schwer B. Cell. 2013; 152: 417–429. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Chiarle R, Zhang Y, Frock RL, Lewis SM, Molinie B, Ho YJ, Myers DR, Choi VW, Compagno M, Malkin DJ, et al. Клетка. 2011; 147: 107–119. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Choi NM, Loguercio S, Verma-Gaur J, Degner SC, Torkamani A, Su AI, Oltz EM, Artyomov M, Feeney AJ.J Immunol. 2013; 191: 2393–2402. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- de Laat W., Grosveld F. Curr Opin Genet Dev. 2007. 17: 456–464. [PubMed] [Google Scholar]
- Frock RL, Hu J, Meyers RM, Ho YJ, Kii E, Alt FW. Nat Biotechnol. 2015; 33: 179–186. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Gostissa M, Schwer B, Chang A, Dong J, Meyers RM, Marecki GT, Choi VW, Chiarle R, Zarrin AA, Alt FW. Proc Natl Acad Sci U S. A. 2014; 111: 2644–2649. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Gostissa M, Yan CT, Bianco JM, Cogne M, Pinaud E, Alt FW.Природа. 2009; 462: 803–807. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Gramlich HS, Reisbig T, Schatz DG. PLoS One. 2012; 7: e39601. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Guo C, Yoon HS, Franklin A, Jain S, Ebert A, Cheng HL, Hansen E, Despo O, Bossen C, Vettermann C, et al. Природа. 2011; 477: 424–430. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Hakim O, Resch W., Yamane A, Klein I, Kieffer-Kwon KR, Jankovic M, Oliveira T, Bothmer A, Voss TC, Ansarah-Sobrinho C и др. .Природа. 2012; 484: 69–74. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Jhunjhunwala S, van Zelm MC, Peak MM, Cutchin S, Riblet R, van Dongen JJ, Grosveld FG, Knoch TA, Murre C. Cell. 2008. 133: 265–279. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Кляйн И.А., Реш В., Янкович М., Оливейра Т., Яман А., Накахаши Н., Ди Вирджилио М., Ботмер А., Нуссенцвейг А., Роббиани Д. Ф. и др. Клетка. 2011; 147: 95–106. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Ковальчук А.Л., Ансара-Собриньо К., Хаким О., Реш В., Толарова Н., Дюбуа В., Яман А., Такидзава М., Кляйн И., Хагер Г.Л. и др.Proc Natl Acad Sci U S. A. 2012; 109: 10972–10977. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Либерман-Эйден Э., ван Беркум Н.Л., Уильямс Л., Имакаев М., Рагоци Т., Теллинг А., Амит И., Ладжой Б.Р., Сабо П.Дж., Доршнер М.О. и др. Наука. 2009. 326: 289–293. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Lin SG, Guo C, Su A, Zhang Y, Alt FW. Proc Natl Acad Sci U S. A. 2015; 112: 1815–1820. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Osborne CS, Chakalova L, Mitchell JA, Horton A, Wood AL, Bolland DJ, Corcoran AE, Fraser P.PLoS Biol. 2007; 5: e192. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Rao SS, Huntley MH, Durand NC, Stamenova EK, Bochkov ID, Robinson JT, Sanborn AL, Machol I, Omer AD, Lander ES, et al. Клетка. 2014; 159: 1665–1680. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Subrahmanyam R, Sen R. Curr Top Microbiol Immunol. 2012; 356: 39–63. [PubMed] [Google Scholar]
- Ван Дж. Х., Гостисса М., Ян К. Т., Гофф П., Хикернелл Т., Хансен Э., Дифилиппантонио С., Веземанн Д. Р., Заррин А. А., Раевски К. и др.Природа. 2009; 460: 231–236. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Заррин А.А., Дель Веккио С., Ценг Э., Глисон М., Зарин П., Тиан М., Альт Ф.В. Наука. 2007; 315: 377–381. [PubMed] [Google Scholar]
Сложные ландшафты соматической перестройки в геномах рака груди человека
Кай, Ф. Дж. Мутации-ассоциированные гены слитного рака в солидных опухолях. Mol. Рак Тер. 8 , 1399–1408 (2009)
CAS Статья Google ученый
Мительман Ф., Йоханссон, Б. и Мертенс, Ф. Слияние генов и реаранжированных генов как линейная функция хромосомных аберраций при раке. Nature Genet. 36 , 331–334 (2004)
CAS Статья Google ученый
Мительман, Ф., Йоханссон, Б. и Мертенс, Ф. Влияние транслокаций и слияния генов на причинность рака. Nature Rev. Cancer 7 , 233–245 (2007)
CAS Статья Google ученый
Futreal, P.A. et al. Перепись генов рака человека. Nature Rev. Cancer 4 , 177–183 (2004)
CAS Статья Google ученый
Страттон, М. Р., Кэмпбелл, П. Дж. И Футреал, П. А. Геном рака. Nature 458 , 719–724 (2009)
CAS Статья ОБЪЯВЛЕНИЯ Google ученый
Сойерс, К. Л. Хронический миелоидный лейкоз. N. Engl. J. Med. 340 , 1330–1340 (1999)
CAS Статья Google ученый
Tomlins, S.A. et al. Рекуррентное слияние генов транскрипционных факторов TMPRSS2 и ETS при раке простаты. Наука 310 , 644–648 (2005)
CAS Статья ОБЪЯВЛЕНИЯ Google ученый
Сода, М.и другие. Идентификация трансформирующего слитого гена EML4 — ALK при немелкоклеточном раке легкого. Nature 448 , 561–566 (2007)
CAS Статья ОБЪЯВЛЕНИЯ Google ученый
Хёглунд, М., Гиссельссон, Д., Салл, Т. и Мительман, Ф. Справляться со сложностями. многомерный анализ кариотипов опухолей. Cancer Genet. Cytogenet. 135 , 103–109 (2002)
Артикул Google ученый
Бигнелл, Г.R. et al. Архитектура соматической перестройки генома в ампликонах рака человека при разрешении на уровне последовательностей. Genome Res. 17 , 1296–1303 (2007)
CAS Статья Google ученый
Volik, S. et al. Профилирование конечных последовательностей: анализ аберрантных геномов на основе последовательностей. Proc. Natl Acad. Sci. США 100 , 7696–7701 (2003)
Артикул ОБЪЯВЛЕНИЯ Google ученый
Руань, Ю.и другие. Транскрипты слияния и транскрибированные ретротранспозированные локусы обнаружены посредством всестороннего анализа транскриптома с использованием парных концевых тегов (ПЭТ). Genome Res. 17 , 828–838 (2007)
CAS Статья Google ученый
Greenman, C. et al. Паттерны соматических мутаций в геномах рака человека. Nature 446 , 153–158 (2007)
CAS Статья ОБЪЯВЛЕНИЯ Google ученый
Дерево, Л.D. et al. Геномные пейзажи рака груди и колоректального рака человека. Наука 318 , 1108–1113 (2007)
CAS Статья ОБЪЯВЛЕНИЯ Google ученый
Исследовательская сеть Атласа генома рака. Всесторонняя геномная характеристика определяет гены и основные пути глиобластомы человека. Nature 455 , 1061–1068 (2008)
Артикул Google ученый
Уир, Б.A. et al. Характеристика генома рака при аденокарциноме легкого. Nature 450 , 893–898 (2007)
CAS Статья ОБЪЯВЛЕНИЯ Google ученый
Кэмпбелл, П. Дж. И др. Идентификация соматически приобретенных перестроек при раке с использованием массового параллельного секвенирования парных концов всего генома. Nature Genet. 40 , 722–729 (2008)
CAS Статья Google ученый
Weterings, E.И Чен, Д. Дж. Бесконечная сказка о негомологичном соединении концов. Cell Res. 18 , 114–124 (2008)
CAS Статья Google ученый
ван Гент, Д. К. и ван дер Бург, М. Негомологичное соединение концов, сложное дело. Онкоген 26 , 7731–7740 (2007)
CAS Статья Google ученый
Хефферин, М.Л. и Томкинсон, А. Е. Механизм репарации двухцепочечных разрывов ДНК путем негомологичного соединения концов. Восстановление ДНК 4 , 639–648 (2005)
CAS Статья Google ученый
Гастингс, П. Дж., Лупски, Дж. Р., Розенберг, С. М. и Ира, Г. Механизмы изменения числа копий гена. Nature Rev. Genet. 10 , 551–564 (2009)
CAS Статья Google ученый
Ян, К.T. et al. Переключение классов IgH и транслокации используют надежный неклассический путь присоединения концов. Nature 449 , 478–482 (2007)
CAS Статья ОБЪЯВЛЕНИЯ Google ученый
Боландер, С. К. ETV6: универсальный участник лейкемогенеза. Семин. Cancer Biol. 15 , 162–174 (2005)
CAS Статья Google ученый
Кнезевич, С.Р., Макфадден, Д. Э., Тао, В., Лим, Дж. Ф. и Соренсен, П. Х. Новое слияние гена ETV6 — NTRK3 при врожденной фибросаркоме. Nature Genet. 18 , 184–187 (1998)
CAS Статья Google ученый
Ланнон, К. Л. и Соренсен, П. Х. ETV6 – NTRK3: химерная протеинтирозинкиназа с активностью трансформации во многих клеточных линиях. Семин. Cancer Biol. 15 , 215–223 (2005)
CAS Статья Google ученый
Джонс, Д.T. et al. Тандемная дупликация, дающая новый онкогенный гибридный ген BRAF , определяет большинство пилоцитарных астроцитом. Cancer Res. 68 , 8673–8677 (2008)
CAS Статья Google ученый
Басеке, Дж., Уилан, Дж. Т., Гризингер, Ф. и Бертран, Ф. Э. Частичная тандемная дупликация MLL при остром миелоидном лейкозе. Br. J. Haematol. 135 , 438–449 (2006)
Артикул Google ученый
Блоу, Дж.Дж. И Гиллеспи, П. Дж. Лицензирование репликации и рак — смертельная путаница? Nature Rev. Cancer 8 , 799–806 (2008)
CAS Статья Google ученый
Perou, C.M. et al. Молекулярные портреты опухолей груди человека. Nature 406 , 747–752 (2000)
CAS Статья ОБЪЯВЛЕНИЯ Google ученый
Сорли Т.и другие. Паттерны экспрессии генов карциномы молочной железы различают подклассы опухолей с клиническими последствиями. Proc. Natl Acad. Sci. США 98 , 10869–10874 (2001)
CAS Статья ОБЪЯВЛЕНИЯ Google ученый
Chin, K. et al. Геномные и транскрипционные аберрации, связанные с патофизиологией рака груди. Раковая клетка 10 , 529–541 (2006)
CAS Статья Google ученый
Бергамаски, А.и другие. Отчетливые закономерности изменения числа копий ДНК связаны с различными клинико-патологическими особенностями и подтипами экспрессии генов рака груди. Genes Chromosom. Рак 45 , 1033–1040 (2006)
CAS Статья Google ученый
Ли, Х., Руан, Дж. И Дурбин, Р. Картирование считываний коротких последовательностей ДНК и вызова вариантов с использованием показателей качества картирования. Genome Res. 18 , 1851–1858 (2008)
CAS Статья Google ученый
Нин, З., Кокс, А. Дж. И Малликин, Дж. С. SSAHA: быстрый метод поиска для больших баз данных ДНК. Genome Res. 11 , 1725–1729 (2001)
CAS Статья Google ученый
Venkatraman, E. & Olshen, A. Более быстрый алгоритм круговой двоичной сегментации для анализа массивов данных CGH. Биоинформатика 23 , 657–663 (2007)
CAS Статья Google ученый
Ламброс, М.и другие. Разблокирование архивов патологий для молекулярно-генетических исследований: надежный метод создания зондов для хромогенной и флуоресцентной гибридизации in situ . Lab. Инвестировать. 86 , 398–408 (2006)
CAS Статья Google ученый
Abeysinghe, S. S. et al. Контрольные точки транслокации и грубой делеции при наследственном заболевании и раке человека I: нуклеотидный состав и мотивы, связанные с рекомбинацией. Hum. Мутат. 22 , 229–244 (2003)
CAS Статья Google ученый
Scholz, F. W. & Stephens, M. A. Тесты Андерсона-Дарлинга с образцом K. Am. J. Stat. Доц. 82 , 918–924 (1987)
MathSciNet Google ученый
Котел эволюции: дупликация, делеция и перестройка в геномах мыши и человека
Реферат
В этом исследовании изучаются геномные дупликации, делеции и перестройки, которые произошли в масштабах от одного основания до полных хромосом, путем сравнения геномов мыши и человека.Из выравнивания полногеномных последовательностей очевидно 344 больших (> 100 т.п.н.) блоков консервативной синтении, но они далее фрагментируются эволюционными событиями меньшего масштаба. Исключая вставки транспозонов, в среднем в каждой мегабазе геномного выравнивания мы наблюдаем две инверсии, 17 дупликаций (пять тандемных или почти тандемных), семь транспозиций и 200 делеций по 100 и более оснований. Это включает 160 инверсий и 75 дупликаций или транспозиций длиной> 100 т.п.н. Частоты этих меньших событий не намного выше в готовых частях сборки.Многие из более мелких транспозиций являются обработанными псевдогенами; мы определяем «синтеническое» подмножество выравниваний, которое исключает эти и другие мелкомасштабные транспозиции. Эти сопоставления свидетельствуют о том, что ≈2% генов общего предка человека / мыши были удалены или частично удалены у мыши. Также оказалось, что у мышей немного меньше индуцированной транспозоном дупликации генома, чем у человека. Хотя некоторые из обнаруживаемых нами событий, возможно, связаны с неправильной сборкой или отсутствием данных в текущей последовательности генома или с ограничениями наших методов, скорее всего, они представляют собой подлинные эволюционные события.Чтобы сделать эти наблюдения, мы разработали новые методы выравнивания, которые могут надежно обрабатывать большие пробелы и различать ортологичные и паралогичные выравнивания.
Evolution создает новые формы и функции в результате взаимодействия воспроизводства, вариаций и отбора. Есть много типов вариаций; наиболее распространенным и хорошо изученным является замена одного основания другим. Небольшие вставки и удаления также довольно распространены. Крупномасштабные вставки обычно включают дублирование части генома.Эти дупликации могут быть отправной точкой для развития нового гена с новой функцией. Эволюция недуплицированных генов обычно сильно ограничивается отбором, потому что существующая функция гена должна поддерживаться. После дублирования одна копия может потерять свою первоначальную функцию и, возможно, принять новую функцию (1, 2). Делеция и перестройка также играют важную роль в долгосрочной эволюции геномов.
В этом исследовании изучаются закономерности вариации, наблюдаемые на всех уровнях, путем сравнения геномов человека и мыши друг с другом.Человек и мышь находятся на отличном расстоянии для изучения всех типов вариаций. Геномы все еще достаточно похожи, чтобы можно было выровнять большую часть ортологичных последовательностей на уровне ДНК (3), но достаточно далеко, чтобы у них была возможность накопиться.
Хромосомные перестройки ≥1 мегабазы можно наблюдать путем сравнения генетических карт организмов (4) и окраски хромосом (5). Приблизительно 200 консервативных блоков синтении между человеком и мышью были обнаружены путем сравнения порядка генов до того, как стали доступны последовательности генома, с недавними оценками в диапазоне от 98 (6) до 529 блоков (7), в зависимости от деталей определения и метода.Было обнаружено, что распределение блоков синтении по длине согласуется с теорией случайного разрушения, введенной Надо и Тейлором (8, 9) до того, как стали доступны важные данные о порядке генов. В недавних сравнениях геномов человека и мыши реаранжировки ≥100000 оснований были изучены путем сравнения 558000 высококонсервативных сравнений коротких последовательностей (средняя длина 340 п.н.) в пределах окна размером 300 т.п.н. Было обнаружено примерно 217 блоков консервативной синтении, образованных из 342 консервативных сегментов, с распределением длины, примерно соответствующим модели случайного разрыва (3).Последующий анализ этих данных обнаружил 281 консервативный блок синтении размером по крайней мере 1 мегабазу с несколькими тысячами дополнительных «микроорганизмов» внутри этих блоков, примерно по одному на мегабазу (10).
Наиболее распространенными вариантами являются одноосновные переходы, то есть замены C / T и G / A (11, 12). Одноосновные вставки и делеции также довольно распространены, хотя они быстро выбираются из кодирующих областей. Замены и небольшие (<20 оснований) вставки и делеции могут быть изучены при традиционном выравнивании нуклеотидов гомологичных геномных последовательностей.Традиционное попарное выравнивание состоит из двух сегментов геномной ДНК с пробелами, помещенными для максимального увеличения количества совпадающих оснований. Простой пример:
Если две последовательности получены от общего предка, то несоответствие может возникнуть в результате замены в любой последовательности относительно их общего предка. Точно так же пробел в выравнивании может быть вызван либо вставкой в одну последовательность, либо делецией в другой.
В основе процесса попарного выравнивания лежит функция подсчета очков, которая присваивает положительные значения совпадающим нуклеотидам и отрицательные значения несовпадениям и пробелам.В большинстве современных программ используется так называемая «аффинная» оценка разрыва, при которой первый символ разрыва в разрыве требует значительных затрат «открытия разрыва», а каждый последующий символ разрыва влечет за собой несколько меньшую стоимость «расширения разрыва». Поскольку промежутки часто имеют длину более одного основания, схемы аффинной оценки моделируют лежащие в основе биологические процессы намного лучше, чем системы оценки с фиксированными промежутками. Показатели аффинных пробелов обычно достаточно хорошо работают для выравнивания белков, где пробелы редки и, как правило, короткие, но также не отражают частоту более длинных пробелов (13-15).Выравнивания нуклеотидов, особенно вне кодирующих областей, как правило, требуют гораздо большего количества пробелов, чем выравнивания белков, и некоторые из пробелов могут быть слишком большими, чтобы их можно было обнаружить с помощью традиционных программ попарного выравнивания (16). Более того, в традиционных программах попарного выравнивания в любом данном месте разрыв может возникать только в одной последовательности. Независимые события делеции у каждого вида, которые удаляют часть, но не всю одну и ту же предковую последовательность, не могут быть представлены, хотя они довольно распространены. В этих случаях программа выравнивания либо разбивает выравнивание на две части, либо заставляет негомологичные основания выравниваться.Традиционные программы также не могут учитывать инверсии, транслокации или дупликации. Они могут выровнять только более короткие сегменты геномной ДНК, в которых не произошло ни одного из этих событий. Следовательно, в то время как значительный анализ вариаций на одной базовой шкале доступен из традиционных выравниваний последовательностей, чтобы дополнить анализ крупномасштабных перестроек, анализ вариации на средних шкалах все еще остается интересной задачей. Роль сегментарного дублирования от нескольких тысяч до нескольких миллионов оснований в нашей собственной эволюции (17) — одно из указаний на важность вариаций на средних масштабах.
В этой статье мы описываем автоматизированные методы объединения традиционных выравниваний в более крупные структуры, цепочки и сети, которые могут эффективно преодолеть разрыв между окраской хромосом и выравниванием последовательностей (рис. 1). Эти методы могут учитывать инверсии, транслокации, дупликации, крупномасштабные делеции и перекрывающиеся делеции. Мы применяем эти инструменты к сгенерированным blastz выравниваниям (18), чтобы исследовать паттерны вариаций, которые произошли во всех масштабах с момента расхождения мышиного и человеческого родов.
Рис.1.Выравнивание мыши / человека по актинину α-3 до и после связывания и связывания, как показано в браузере генома на http://genome.ucsc.edu. Дорожка RefSeq генов показывает структуру экзона / интрона этого человеческого гена, который имеет ортолог, а также несколько паралогов и псевдогенов у мыши. Дорожка мыши all blastz показывает выравнивание blastz, окрашенное хромосомой мыши. Ортологичный ген находится на хромосоме 19 мыши, которая окрашена в фиолетовый цвет. Хотя blastz находит гомологию очень чувствительным образом, она фрагментирована.Дорожка chained blastz показывает выравнивания после связывания. Цепочка связывает связанные фрагменты. Ортологичные гены и паралоги представляют собой единое целое. Цепочка также объединяет некоторые избыточные выравнивания и устраняет несколько изолированных выравниваний с очень низкой оценкой. Дорожка Mouse / Human Alignment Net предназначена для отображения только ортологичных выравниваний. В этом случае не было никаких перестановок, кроме вставок и удалений среднего размера, поэтому чистый трек довольно прост. Нажатие на цепочку или сетевую дорожку позволяет пользователю открыть новый браузер в соответствующем регионе другого вида.
Методы
Замораживание генома человека в ноябре 2002 г. и замораживание генома мыши в феврале 2002 г. были взяты с http://genome.ucsc.edu. Два генома были выровнены программой blastz, как описано в ссылке. 18, за исключением того, что в дополнение к маскированию повторов транспозонов [идентифицированных с помощью repeatmasker (19)], простые повторы периода 12 или меньше, обнаруженные тандемным поиском повторов (20), были замаскированы, а последовательность мыши «chrUn», склонная к артефактам, последовательность, которая не могла быть собрана в большие контиги или эффективно картирована в хромосомах мыши, была исключена.Выравнивания blastz были преобразованы в формат axt (см. Ссылку 18) с помощью программы lavtoaxt.
Используя ту же матрицу подсчета нуклеотидов, что и у blastz, но новую схему подсчета с кусочно-линейными пробелами, новая программа axtchain сформировала максимально подсчитывающую цепочку выравниваний из непрерывных частей входных выравниваний (рис. 1). Цепное выравнивание (или «цепочка») между двумя видами состоит из упорядоченной последовательности традиционных попарных выравниваний нуклеотидов («блоков»), разделенных большими пробелами, некоторые из которых могут быть одновременными пробелами у обоих видов.Порядок блоков в цепи должен соответствовать порядку геномной последовательности у обоих видов. Таким образом, цепь не может иметь локальных инверсий, транслокаций или дупликаций среди частей ДНК, которые она выравнивает. Однако цепочке разрешено пропускать сегменты ДНК у одного или обоих видов. В частности, вмешивающаяся ДНК одного вида, которая не совпадает с другой, потому что она локально инвертирована или была вставлена посредством клон-специфической транслокации или дупликации, пропускается во время построения цепи.Таким образом, цепь может представлять собой широко разбросанные фрагменты геномной ДНК двух современных видов, которые фактически произошли от одного геномного сегмента общего предка без перегруппировки.
Для эффективного построения цепочек axtchain использует вариант алгоритма на основе k-мерного дерева (kd-tree), описанного в исх. 21. Для обнаружения случаев перекрывающихся делеций у обоих видов подсчет определяется таким образом, чтобы программа выравнивания обычно открывала одновременный пробел, а не вынуждала выравнивать негомологичные области, потому что штрафы, понесенные за серию несоответствий, превышают штраф за одновременный пробел.Эта стратегия разрешения одновременных пропусков также помогает справиться с локальными инверсиями и отсутствующими последовательностями (блоками Ns) в незавершенных геномах. Результирующие цепочки часто охватывают несколько мегабаз.
Для многих целей нужно выбрать только одно лучшее выравнивание для каждой области генома человека. Ранее (18) для этой цели мы разработали программу axtbest. Однако цепи во многих отношениях представляют собой лучший субстрат для выбора «лучшего» выравнивания, чем одиночные выравнивания blastz.Мы разработали новую программу, chainnet, чтобы улучшить этот процесс. В этой программе все основания во всех хромосомах изначально помечаются как неиспользуемые. Затем цепочки помещаются в список, отсортированный по цепочке с наибольшим количеством очков. Программа переходит в цикл, на каждой итерации удаляя следующую цепочку из списка, отбрасывая части цепочки, которые пересекаются с основаниями, уже покрытыми ранее взятыми цепями, а затем помечают базы, которые остались в цепочке, как покрытые. . Программа использует красно-черные деревья, чтобы отслеживать, какие области хромосомы уже покрыты.Если цепочка покрывает базы, которые находятся в промежутке в ранее взятой цепочке, она помечается как дочерняя по отношению к предыдущей цепочке. Таким образом образуется иерархия цепочек, которую мы называем сетью. Программа chainnet также отслеживает, какие основания покрываются более чем одной цепочкой, чтобы помочь отличить дублированные области от недуплицированных.
Полученные сетевые файлы дополнительно аннотируются программой netsynteny, которая отмечает, какие цепи в сети инвертированы относительно своих родителей, смещены или находятся на разных хромосомах.Дополнительные аннотации к повторам, включая повторы, специфичные для клонов, и повторы, предшествующие разделению мыши / человека, добавляются программой netclass. Короткие цепи, встроенные в более длинную цепь, которая происходит из областей, отличных от области более длинной цепи, а также короткие цепи между длинными цепями на верхнем уровне сети часто являются процессируемыми псевдогенами. Поскольку это может затруднить анализ многих типов, мы также подготовили подмножество цепочек, которые считаются «синтенными». Чтобы считаться синтенической, цепь должна либо иметь очень высокий балл, либо быть встроенной в большую цепь на той же хромосоме и происходить из того же региона, что и большая цепь.Таким образом, инверсии и тандемные дупликации считаются синтеническими. Синтеническое подмножество выравниваний было создано с помощью программы netfilter с использованием флага –sin. Хотя netfilter не устраняет псевдогены, возникающие в результате тандемного дублирования, он устраняет обработанные псевдогены, которых гораздо больше.
Дополнительные сведения об этих алгоритмах описаны в исходном коде, который, наряду с исполняемыми файлами Linux для lavtoaxt, axtchain, chainnet, netsynteny, netclass и netfilter, доступен по адресу www.soe.ucsc.edu/~kent. Цепочки и сети отображаются вместе с другими аннотациями в браузере генома по адресу http://genome.ucsc.edu (22), а также могут быть загружены массово с этого сайта.
Результаты
Первоначальные выравнивания мышей blastz покрывают 35,9% генома человека. Это меньше, чем 39,9%, о которых говорится в исх. 3, из-за маскирования тандемных повторов периода 12 и менее и удаления склонной к артефакту мышиной последовательности в «ChrUn». Построение более длинных цепочек из начальных выравниваний blastz привело к меньшему количеству и существенно более длинным выравниваниям (рис.1). Длину связанного выравнивания можно измерить двумя разными способами. Мы определяем (человеческий) отрезок цепи как расстояние в основаниях в геноме человека от первого до последнего человеческого основания в цепи, включая промежутки, и мы определяем размер цепи как количество выравнивающих оснований в это, не считая пробелов. Оба они показали существенное увеличение за счет построения цепочки, при этом средний размах человека увеличился с 608 до 22 830 п.н., а средний размер увеличился с 574 до 7 062 п.н.Поскольку многие более мелкие выравнивания blastz часто объединялись в одну более длинную цепочку, также наблюдалось существенное сокращение общего числа выравниваний, задействованных в сравнении человека и мыши, с ≈8,5 миллиона до ≈150 000. В некоторых случаях небольшие выравнивания blastz с низким показателем также отбрасываются после объединения в цепочку. Это приводит к уменьшению с 35,9% до 34,6% оснований в геноме человека, сопоставленных с мышиным. Эти и другие сравнительные статистические данные приведены в таблице 1.
Таблица 1. Сравнение выравниваний / цепочек blastz до и после обработки с помощью axtchain и после построения человеческой сетиПолногеномная коллекция этих цепочек была использована для изучения распределения длин промежутков, вызванных инделками всех размеров, начиная с расхождения этих видов, от единичных оснований до инделек размером в несколько килобайт.Модель оценки аффинных пробелов, обычно используемая в программах выравнивания последовательностей, соответствует статистической модели, в которой размеры отступов следуют геометрическому распределению, в котором количество пробелов размером N + 1 будет постоянной долей количества пробелов размером N для всех N. Наблюдаемое нами распределение длины нарушает эту модель. На рис. 2 показана гистограмма размеров зазоров, наблюдаемых в наборе построенных цепочек, с частотой появления для различных длин отступов, нанесенной в логарифмическом масштабе.На таком графике геометрическое распределение выглядело бы как прямая линия. На самом деле сюжет довольно сложный. В общем, существует больше коротких и длинных промежутков, чем мы могли бы видеть в геометрическом распределении, и есть резкие всплески в количестве промежутков, наблюдаемых в человеческой последовательности на ≈300 оснований, соответствующих вставкам ALU (19).
Рис.2.( a ) Небольшие вставки и удаления. ( b ) Большие вставки, делеции и транспозоны. Графики числа вставок и делеций по всему геному в сравнении сих размер. Синяя линия показывает комбинацию человеческих вставок и делеций мыши, тогда как красная линия показывает мышиные вставки и человеческие делеции. Вертикальная шкала — это натуральный логарифм количества вставок / удалений такого размера. В b подсчеты сгруппированы в ячейки по 25. Зеленая линия показывает процент оснований в промежутках мыши такого размера, которые покрыты специфичными для человека транспозонами. Пики на графиках вставки / удаления, по-видимому, связаны с транспозонами.
Цепи также содержали значительное количество одновременных разрывов у обоих видов (рис. 3). Для небольших зазоров одновременные зазоры довольно редки, но это явление становится все более важным с увеличением размера зазора. Частота появления больших одновременных промежутков примерно соответствует модели, в которой они возникают из двух независимых событий indel, по одному для каждого вида (рис. 3 c ).
Рис.3.( a ) Лог-гистограмма частот пропусков для пропусков длиной <50 базисов.( b ) Лог-гистограмма частот пропусков для пропусков длиной до 1500 базисов. ( c ) Регистрируйте гистограмму частот пропусков для пропусков длиной до 50 000 базисов. Относительная частота одновременных и одиночных пропусков показана в обеих последовательностях. Горизонтальная ось используется для пробелов в последовательности мыши, которые представляют либо вставки у человека, либо делеции у мыши. Вертикальная ось используется для зазоров в человеческом теле. Журнал количества одновременных пропусков определенного диапазона размеров преобразуется в уровень серого для создания двухмерной гистограммы.Горизонтальная и вертикальная оси не прорисовываются; темные линии на оси должны отражать логарифмические частоты пропусков, которые находятся только в одной последовательности. ( a ) Разрывы <50 баз. Здесь особенно заметны пробелы, которые есть чисто у мыши или чисто у человека. ( b ) Пробелы 10–1 500 баз. Эффекты, индуцированные транспозоном на рис. 1, также можно увидеть здесь. Обратите также внимание на относительную концентрацию около диагонали для вставок <200 оснований. Это происходит в основном из-за небольших инверсий и локально расходящейся последовательности.( c ) Пробелы в 1000–50 000 баз. В этом диапазоне логарифмическая частота одновременных пропусков данного комбинированного (человека и мыши) размера зазора отличается примерно на константу от суммы логарифмических частот отдельных односторонних размеров зазора у каждого вида. В этом смысле эти более длинные одновременные промежутки действуют так, как будто они возникают из независимых промежутков в каждом отдельном виде.
Вложенная «сетевая» структура цепей, созданная программой chainnet, была использована для исследования перестроек, которые произошли после расхождения человека и мыши в форме инверсии (рис.4), делеции, транспозиции, ретротранспозиции, тандемной дупликации и перемежающейся дупликации. Из-за различных типов дупликаций 52% кодирующих областей и 3,3% генома человека в целом охвачены более чем одним выравниванием мыши blastz (3) даже после исключения транспозонов. Это может затруднить отделение ортолога от паралога и гена от псевдогена. Структура сети помогает устранить их неоднозначность. Структура сети не является симметричной между видами: человеческая сеть конструируется, позволяя только единственной наилучшей ДНК из генома мыши выровняться с любым отдельным местом в геноме человека, а сеть мыши строится противоположным образом.Это одностороннее требование «наилучшего в геноме» не так строго, как требование «взаимного наилучшего соответствия», когда разрешены только сопоставления, наилучшие в геноме для обоих видов. Последний запрещает исследование дупликаций, специфичных для клонов, путем межвидового выравнивания, потому что он позволяет выровнять не более одной копии дублированной области с другим видом, и часто даже одна копия не может быть полностью выровнена. У человека и мыши это приводит к значительному сокращению общего количества геномной ДНК, охватываемой межвидовым выравниванием.Мы обнаружили, что требование взаимного наилучшего выравнивания снижает охват в сети для людей на 11%, а в сети для мышей — на 9%. Эти числа довольно близки, что позволяет предположить, что одинаковый уровень дупликации наблюдается в обоих геномах со времен общего предка. В дальнейшем мы продолжим исследовать человеческую сеть, построенную, как описано в Методы , без требования взаимного наилучшего.
Рис.4.Инверсия из 15 000 оснований, содержащая два транскрипта и показывающая chr7: 2077222–2497100 в сборке генома человека в ноябре 2002 года.Многочисленные более мелкие перестановки также видны на чистом треке на этом рисунке. В некоторых случаях меньшие из них просто представляют собой паралогичные мышиные области, которые заполняются, когда ортологичная мышиная область еще не секвенирована.
На основе анализа человеческой сети частота различных реаранжировок на черновой последовательности мышей в целом и на 48-миллионном (2%) подмножестве завершенной последовательности показана в таблице 2. В целом паттерны реаранжировки были очень похожи в законченном подмножестве генома мыши, как и в геноме в целом.То же самое относится и к готовой подгруппе генома человека (данные не показаны). В готовом подмножестве было значительно больше локальных дупликаций, что, вероятно, отражало исчезновение локальных дупликаций, общий артефакт полногеномных методов сборки дробовика, используемых у мышей. В общем, наиболее распространенный тип реаранжировки — это дублируемый участок генома, который вставляется в другую хромосому (несинтетическая дупликация). Большинство из них, по-видимому, являются обработанными псевдогенами. На удивление часто встречались несинтетические видимые транспозиции.
Таблица 2. Статистика реаранжировки генома мыши относительно генома человекаРаспределение пролетов цепей из человеческой сети (рис. 5) показывает примерно бимодальное распределение для цепей, которые охватывают <100 000 оснований («короткие цепочки»), и длинный плоский хвост, состоящий из 579 цепей, охватывающих от 100 000 оснований. и ≈115 миллионов оснований («длинные цепочки», средняя длина 983 кб). В совокупности длинные цепи охватывают 90,9% генома человека (исключая пробелы <100000 оснований в отдельных цепях), а их выровненные основания покрывают 32.9% оснований в геноме человека. Напротив, все цепи вместе (включая произвольно большие промежутки в отдельных цепях) охватывают 96,3% генома человека и соответствуют 34,6% его. Таким образом, одни только длинные цепи (без их длинных промежутков) охватывают 94,4% оснований, охватываемых всеми цепями, и включают 95,1% всех выровненных оснований.
Рис.5.Распределение пролётов всех 147 445 цепей в человеческой сети. Распределение состоит из бимодального участка для коротких цепочек с размахом <10 5 и плоского хвоста для 579 длинных цепочек размером от 10 5 до ≈10 8 .
Из 579 длинных цепочек 344 появляются на верхнем уровне сети и образуют большие первичные единицы синтении с мышью. Расположение этих цепей в значительной степени согласуется с областями синтении у мышей, идентифицированными в более ранних исследованиях (см. Обсуждение , ). Остальные 235 длинных цепей появляются на более низких уровнях сети, потому что они встроены в промежутки этих первичных цепочек синтении. Из них 160 представляют собой инверсии, 29 представляют собой дупликации или транслокации соседних областей на одной и той же хромосоме, а 46 представляют собой удаленные дупликации или транслокации на большом расстоянии в одной и той же хромосоме или между хромосомами.Некоторые из них также были обнаружены в более ранних исследованиях.
В дополнение к 344 длинным цепочкам на верхнем уровне сети существует 19 800 коротких цепочек на верхнем уровне и намного больше на нижних уровнях. Короткие цепи на верхнем уровне часто появляются в длинных сериях в регионах, где ранее не было обнаружено значительной синтении между геномами человека и мыши, и представляют собой то, что кажется горячими точками для перегруппировок или дупликаций. В этих регионах лучшее выравнивание у других видов быстро смещается от одной хромосомы к другой.Многие из этих регионов содержат кластеры генов из семейств, которые недавно подверглись клоноспецифической экспансии. Они включают множество генов, участвующих в иммунной системе, обонятельных рецепторах (23) и цинковых пальцах типа Krüppel-бокс C2h3-типа, которые, как известно, сильно дублировались и были мобильны в эволюции млекопитающих (24). Таблицу таких регионов см. В Таблице 3, которая опубликована в качестве вспомогательной информации на веб-сайте PNAS www.pnas.org.
Поскольку 344 большие цепи, которые находятся на верхнем уровне сети, подобны областям синтении, идентифицированным в более ранних исследованиях, распределение их промежутков примерно согласуется с простой моделью случайного обрыва, которая хорошо работала в предыдущей синтении (8). .Однако наличие большого количества маленьких цепочек предполагает, что другие процессы могут работать не только внутри, но и между блоками синтении, определяемыми длинными цепочками. Эти промежуточные короткие цепи нельзя объяснить как дополнительные блоки синтении вместе с более длинными блоками, используя параметры простой модели случайного обрыва.
Наконец, как описано в методике , мы также получили «синтеническое» подмножество человеческой сети. Полная сеть покрывает 96.7% оснований в кодирующих областях RefSeq (www.ncbi.nlm.nih.gov/RefSeq; ref. 25), тогда как синтенное подмножество покрывает 93,0%. Полная сеть покрывает 71% оснований в псевдогенах на хромосоме 22, аннотированных Центром Сэнгера (26), тогда как синтеническое подмножество покрывает 13%. Таким образом, синтеническое подмножество должно быть полезно при поиске псевдогенов и в приложениях для анализа генов, где важна специфичность, например, как описано в refs. 27 и 28.
Мы подробно исследовали кодирующие области, покрытые полной сетью, но не синтенической сетью на хромосоме 22.Это включает в себя часть или все 40 из 450 генов RefSeq, картированных в хромосоме 22 в сборке ноября 2002 г. на http://genome.ucsc.edu. Восемнадцать из этих генов находились в областях, которые кажутся горячими точками для перестройки и дупликации, как обсуждалось выше. Двенадцать генов, части которых присутствуют в полной сети, но не в синтенической сети, вызваны пробелами в геноме проекта мыши. В полной сети паралоги заменяются отсутствующими ортологическими регионами. Оказалось, что у мыши были удалены или частично удалены восемь генов.Экстраполяция этого на полный геном будет означать, что мышь удалила или частично удалила ≈2% генов, присутствующих в общем предке. Два из генных картирований RefSeq, отсутствующие в синтенической сети, при ближайшем рассмотрении оказались отображениями на обработанные псевдогены. По мере того, как геномы человека и мыши становятся более законченными, синтенная подсеть будет все более полезной для идентификации обработанных псевдогенов и изучения эволюции генов во всем геноме.
Обсуждение
Представленная здесь техника цепочек и сетей очень полезна для отслеживания эволюции генома.Он имеет несколько основных преимуществ перед такими методами, как GRIMM (4) для изучения геномных перестроек. Он может обрабатывать дублирование и удаление, а также транспонирование и инверсию и обеспечивает разрешение вплоть до базового уровня. Однако метод сетки предполагает, что каждая перегруппировка независима и не перекрывает другие перегруппировки. Он может обнаруживать и правильно классифицировать транслокации, только если они вставляются в середину хромосомы, а не на ее концах. GRIMM может иметь дело с перекрывающимися перестройками и транслокациями на концах хромосом.Когда мы ограничиваем наше внимание блоками из 100 000 оснований или более, мы находим 160 инверсий, что аналогично 149 инверсиям 1 миллиона оснований или более, о которых сообщалось с помощью GRIMM. Уровень транспозиций <100 000 оснований ниже, чем наблюдается в GRIMM, из-за ограничений нашей техники. Точки излома, которые мы наблюдаем между большими синтеническими областями, действительно хорошо коррелируют с теми, которые наблюдаются с GRIMM и с техниками на основе окон, такими как описанные в исх. 3. Сравнение цепей из человеческой сети и блоков синтении, найденных другими методами, можно рассматривать как параллельные треки в браузере генома с помощью ссылки http: // genome.ucsc.edu/cauldron/syntenies.html.
Ясно, что инверсии и другие перестановки происходят на всех масштабах, и действительно, есть больше инверсий <1000 оснований, чем инверсий <1000 оснований. Из-за ограничений в наших методах выравнивания мы, возможно, в настоящее время не наблюдаем многие из мельчайших перегруппировок, особенно те, которые <50 оснований. Наиболее частая перестройка, которую мы наблюдаем (за исключением вставки транспозона), - это несинтеническая дупликация, большая часть которой, по-видимому, происходит из-за процессированных псевдогенов, созданных механизмом транспозона.Однако существует удивительное количество несинтетических недублирующих транспозиций небольших блоков. Некоторые из них, а также другие события, которые мы регистрируем, могут быть артефактами из-за неполноты генома мыши: вторые наилучшие совпадения с геномом мыши иногда используются в сети, где наилучшее совпадение отсутствует, потому что оно попадает в несеквенированный область в геноме мыши. Однако это не объясняет большинство событий. В настоящее время мы не знаем механизма, который генерировал бы движение таких маленьких кусочков генома, как этот.
Наконец, эта работа позволяет нам определить ортологичные гены мыши и человека более четко, чем мы могли в прошлом, и может помочь устранить ложную аннотацию псевдогенов как истинных генов.
Благодарности
Мы благодарим Международный консорциум по секвенированию генома человека и Консорциум по секвенированию генома мышей за предоставленные данные. W.J.K., R.B., A.H. и D.H. были поддержаны Национальным институтом исследования генома человека, грант 1P41HG-02371.D.H. также поддерживал Медицинский институт Говарда Хьюза. W.M. поддерживается грантом HG-02238 Национального института исследования генома человека.
Сноски
↵ † Кому следует направлять корреспонденцию. Электронная почта: kent {at} biology.ucsc.edu.
Этот документ был отправлен напрямую (Трек II) в офис PNAS.
См. Комментарий на странице 11188.
- Авторские права © 2003, Национальная академия наук
Зависимая от вставки последовательности плазмидная перегруппировка у Aeromonas salmonicida вызывает потерю системы секреции третьего типа
Abstract
Aeromonas salmonicida , бактериальный патоген рыб, обладает функциональной системой секреции третьего типа (TTSS), которая необходима для его вирулентности.Гены этой системы в основном расположены в одной области большой плазмиды pAsa5. Бактерии теряют область TTSS из этой плазмиды в результате перестройки при росте в стрессовых условиях роста. Геном A. salmonicida богат последовательностями вставки (IS), которые представляют собой мобильные элементы ДНК, которые могут вызывать перестройки ДНК у других видов бактерий. pAsa5 обладает множеством ИС. Три IS 11 из семейства IS 256 окружают переупорядоченные области.Чтобы подтвердить, что эти IS 11 участвуют в реаранжировках pAsa5, 26 штаммов, происходящих от штамма A449, и два канадских изолята (01-B526 и 01-B516) с реаранжировкой pAsa5 были протестированы с использованием подхода ПЦР, чтобы определить, были ли перегруппировки результат процесса, зависящего от IS 11 . Девять из 26 штаммов дали положительный результат ПЦР, что позволяет предположить, что реаранжировка в этих штаммах была IS-зависимой. Анализ ПЦР показал, что все перегруппировки в штаммах, полученных из A449, были IS 11 -зависимым процессом, в то время как перегруппировки в 01-B526 и 01-B516 могли быть только частично связаны с действием IS 11 .Неидентифицированные элементы, которые влияют на IS-зависимые перегруппировки, могут присутствовать в 01-B526 и 01-B516. Наши результаты показали, что в перегруппировке pAsa5 участвует IS 11 . Это первое исследование, показывающее, что IS участвуют в нестабильности плазмиды у A. salmonicida .
Образец цитирования: Tanaka KH, Dallaire-Dufresne S, Daher RK, Frenette M, Charette SJ (2012) Зависимая от вставки последовательности плазмидная перегруппировка в Aeromonas salmonicida вызывает потерю системы секреции третьего типа.PLoS ONE 7 (3): e33725. https://doi.org/10.1371/journal.pone.0033725
Редактор: Михаэль Хензель, Оснабрюкский университет, Германия
Поступила: 29 сентября 2011 г .; Одобрена: 16 февраля 2012 г .; Опубликовано: 14 марта 2012 г.
Авторские права: © 2012 Tanaka et al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: KHT получил стипендии от Программы присуждения премий студентам бакалавриата по исследованиям в области естественных наук и инженерных исследований Канады (NSERC). SDD получила стипендию фондовой биржи FONCER от Réseau Aquaculture Québec (RAQ) и стипендию от Le Fonds québécois de la recherche sur la nature et les technologies (FQRNT). РКД получил стипендию от Program canadien de bourses de la francophonie (PCBF). Этот проект финансировался грантами RAQ и NSERC.SJC является научным сотрудником Fonds de la Recherche en Santé du Québec (FRSQ). Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили, что конкурирующих интересов не существует.
Введение
Грамотрицательные бактерии Aeromonas salmonicida subsp. salmonicida — этиологический агент фурункулеза, заболевания, которое вызывает сепсис и некроз, особенно у лососевых [1], [2].Это заболевание оказывает серьезное экономическое влияние на рыбоводство.
A. salmonicida обладает широким спектром факторов вирулентности, включая внеклеточные протеазы, липазы и систему секреции функционального типа 3 (TTSS) [3], [4]. TTSS, система вирулентности, разделяемая многими грамотрицательными бактериями, может перемещать эффекторные белки в клетки-хозяева, где они могут нарушать клеточные процессы и нарушать иммунные ответы [5], [6]. TTSS важен для вирулентности A.salmonicida у рыб и Dictyostelium discoideum амеба [7], [8], [9], [10].
В то время как A. salmonicida может содержать множество плазмид, наличие плазмид варьируется в зависимости от штамма. Например, эталонный штамм A449 несет pAsa4, большую плазмиду, которая еще не была четко задокументирована в других штаммах A. salmonicida [11]. В то время как многие штаммы A. salmonicida несут четыре небольшие плазмиды (pAsa1, pAsa2, pAsa3 и pAsal1), в A449 отсутствует pAsal1 [12].Большинство генов, кодирующих TTSS, расположены в одном локусе на pAsa5, за исключением эффекторных белков, кодирующих гены aopP , расположенные на pAsal1, и aexT , расположенные на хромосоме, [11], [13], [14], [15] . Некоторые исследования показали, что pAsa5 является нестабильной плазмидой [10], [13]. Хотя сначала предполагалось, что pAsa5 теряется при выращивании A. salmonicida в стрессовых условиях, например при 25 ° C [13], оказалось, что pAsa5 подвергается перестройке, которая приводит к потере TTSS и, как следствие, , потеря вирулентности [10].
Согласно Daher et al., На pAsa5 происходят два основных вида перестроек, которые приводят к различным профилям потерь [10]. Наиболее частая перестройка приводит к профилю потерь 1-го типа (рис. 1), где теряется только локус TTSS. Менее частая реаранжировка приводит к профилю потери типа 2, когда локус TTSS теряется вместе с областью размером 40 т.п.н., расположенной непосредственно выше по течению (Рис. 1).
Фигура 1. Последовательности вставки (IS 11 s) фланкируют области плазмиды pAsa5 в A.salmonicida , которые потеряны.
Карта линейной плазмиды pAsa5 показывает область TTSS и профили потерь различных штаммов A. salmonicida из таблицы 1, полученные в предыдущем исследовании путем выращивания штаммов при 25 ° C в течение двух недель [10]. Черные прямоугольники обозначают положительные сигналы ПЦР для соответствующих генов, а белые прямоугольники обозначают отрицательные сигналы. Профиль потерь 1-го типа является наиболее распространенным из трех [10]. Стрелки под картой указывают IS на pAsa5.
https: // doi.org / 10.1371 / journal.pone.0033725.g001
Анализ последовательности ДНК pAsa5 выявил присутствие 11 копий инсерционных последовательностей (IS) (Рисунок 1) [11]. Наличие большого количества IS не является исключительной особенностью pAsa5 в A. salmonicida . Геном штамма A449 несет 88 копий различных IS, что больше, чем обычно встречается у γ-протеобактерий [16]. Две копии IS 11 (также называемого IS As3 [11]), члена семейства IS 256 [11], примыкают к TTSS-кодирующей области pAsa5 (рисунок 1).Члены семейства IS 256 были обнаружены как у грамотрицательных, так и у грамположительных эубактерий [17]. Эти мобильные генетические элементы являются частью составного транспозона Tn 4001 [18]. Наличие нескольких копий этих IS связано с гомологичной геномной рекомбинацией [19] и слиянием репликонов [20]. Основываясь на этих выводах, мы предположили, что копии IS 11 на pAsa5 (IS 11 A, IS 11 B и IS 11 C) (Рисунок 1) вносят вклад в перестройки плазмиды, которые приводят к потере TTSS.Таким образом, мы исследовали, участвует ли IS 11 в перестройках pAsa5. Мы использовали подход ПЦР, основанный на праймерах, нацеленных на IS-фланкирующие области, для анализа банка TTSS-дефицитных штаммов.
Материалы и методы
Штаммы бактерий и условия роста
Исходные и производные штаммы A. salmonicida , использованные в этом исследовании, перечислены в таблице 1. Производные штаммы были созданы в предыдущей работе [10]. Вкратце, эти производные были получены путем инокуляции родительских штаммов из замороженных исходных культур на фурункулезном агаре (10 г бакто-триптона, 5 г дрожжевого экстракта, 1 г L-тирозина, 2.5 г NaCl и 15 г агара на литр дистиллированной воды) [21]. Чашки с агаром инкубировали в течение двух недель при 25 ° C. Затем было проведено генотипирование с помощью ПЦР для выявления делеций в pAsa5 [10]. В настоящем исследовании использовались производные, которые потеряли область TTSS (профиль потерь типа 1), и производные, которые также потеряли область выше по течению (профиль потерь типа 2). 01-B526-R4 был сохранен для дальнейшего анализа, даже если он потерял область TTSS, а также другой ген на 40 т.п.н. выше TTSS [10]. Все штаммы инокулировали на фурункулезный агар из замороженных исходных культур и выращивали при 18 ° C в течение двух или трех дней перед лизированием для генотипирования.
Анализ ПЦР
матриц ДНК получали путем лизирования одной бактериальной колонии родительского и производного штаммов в 20 мкл буфера SWL (50 мМ KCl, 10 мМ Трис, pH 8,3, 2,5 мМ MgCl 2 , 0,45% NP-40 и 0,45% Твин 20) [22]. Лизаты нагревали при 95 ° C в течение 5 мин. Смесь для ПЦР содержала 4 мкл 5-кратного буфера Go-Taq (Promega, Мэдисон, Висконсин, США), 1,6 мкл 12,5 мМ dNTP, 1,3 мкл прямого и обратного праймеров (по 100 нг / мкл каждый), 0,1 мкл GoTaq ( 5 У, Промега), 10.7 мкл H 2 O и 1 мкл ДНК-матрицы [22]. Программа ПЦР была следующей: 2 мин 30 с при 95 ° C, 30 циклов по 30 с при 95 ° C, 30 с при 55 ° C и 2 мин при 68 ° C с последующим окончательным продлением на 10 мин при 68 ° С. Образцы разделяли на 1% агарозном геле и окрашивали бромистым этидием 0,5 мкг / мл. ПЦР-анализы проводили не менее двух раз.
Праймеры для ПЦР перечислены в таблице 2. Праймеры, не описанные в литературе, были созданы с использованием программного обеспечения для создания праймеров, доступного в Интернете на веб-сайте Integrated DNA Technologies (www.idtdna.com). Последовательности хромосомы и плазмиды pAsa5 из A. salmonicida A449 и последовательность плазмиды pAsal1 из A. salmonicida JF2267 были использованы для создания праймеров (номера доступа в GenBank: CP000644, CP000646 и AJ508382) [11] , [14].
На протяжении всего исследования было выполненоконтролей ПЦР для подтверждения присутствия ДНК в бактериальных лизатах с использованием пары праймеров, обнаруживающих ген tapA (таблица 2), который переносится хромосомой.Кроме того, чтобы гарантировать, что лизаты, демонстрирующие отсутствие амплификации с праймерами, нацеленными на гены traC , exsD и resD , обнаруженные на pAsa5, не были вызваны ненадлежащей работой праймеров или анализа ПЦР, эти праймеры также тестировали параллельно с лизаты ДНК, которые, как известно, содержат эти гены (обычно лизаты A449). Наконец, все новые праймеры, разработанные в этом исследовании, были протестированы с помощью соответствующего анализа ПЦР, чтобы подтвердить, что они не вызывают неспецифической амплификации отдельно или в комбинации.
Секвенирование ампликона
Положительных продуктов ПЦР, которые подтвердили перестройки B – C или A – C (т.е. продукты размером 2 т.п.н., полученные с парами праймеров 11B1F-11CR и 11AF-11CR, соответственно), очищали с использованием наборов для очистки PureLink PCR (Invitrogen, Carlsbad, CA, USA) и были отправлены на платформу анализа генома (IBIS, Université Laval, QC, Канада) для секвенирования. Был проведен нуклеотидный BLAST-анализ последовательностей, чтобы убедиться, что продукты ПЦР содержат рекомбинированные IS.
Выделение плазмидной ДНК и профили рестрикционных ферментов
Экстрактыплазмид из штаммов, производных 01-B526 и 01-B516, получали с использованием системы очистки ДНК Wizard Plus SV Minipreps (Promega).Экстракты (5 мкл) смешивали с 3,5 мкл H 2 O, 1 мкл 5-кратного реакционного буфера (New England Biolabs, Ипсвич, Массачусетс, США) и 0,5 мкл EcoRI (5U, New England Biolabs). Смеси инкубировали 2 ч при 37 ° C. Образцы ДНК разделяли на 0,7% агарозном геле и окрашивали бромидом этидия (0,5 мкг / мл).
Результаты и обсуждение
Чтобы определить, произошли ли перегруппировки, зависимые от IS 11, в плазмидах pAsa5 производных штаммов, демонстрирующих профили потери pAsa5 типов 1 или 2, необходимо было сконструировать соответствующие праймеры, а перегруппированные IS должны были быть подвергнуты ПЦР. -усиленный.Связанные с IS праймеры были адаптированы для распознавания последовательностей, фланкирующих соответствующий IS (таблица 2). Однако праймеры pAsa5 не были разработаны для получения ампликонов со стандартной конфигурацией плазмиды. Если два идентичных IS на одной плазмиде подвергаются гомологичной рекомбинации, область плазмиды может быть вырезана (рис. 2A). Таким образом, праймеры были сконструированы на основе областей, окружающих IS 11 s. Комбинация этих праймеров (11B1F и 11CR или 11AF и 11CR), таким образом, не будет производить ампликоны при нормальных обстоятельствах, поскольку они находятся слишком далеко друг от друга (рис. 2B).Однако, если pAsa5 подвергается перегруппировке, управляемой процессом, зависящим от IS 11 , праймеры закрываются. Например, для реаранжировки с участием IS 11 B и IS 11 C (перегруппировка B – C) праймеры 11B1F и 11CR будут фланкировать рекомбинированный IS и производить ампликон (рис. 2B), соответствующий профилю потери типа 1. (потеря области ТЦС). Для перегруппировки с участием IS 11 A и IS 11 C (перегруппировка A – C) праймеры 11AF и 11CR будут давать ампликон из-за рекомбинации IS 11 A с IS 11 C.
Рис. 2. IS-зависимая перегруппировка может объяснить профили потерь 1 и 2 и может быть оценена с помощью ПЦР.
A. Два IS могут привести к вырезанию области плазмиды путем гомологичной рекомбинации. B. Это событие, которое может генерировать профиль потерь типа 1 (TTSS) и профиль потерь типа 2 (TTSS и восходящий локус) в pAsa5, можно оценить с помощью ПЦР. На этой карте праймеры (таблица 2), показанные стрелками, фланкируют одну сторону их соответствующих IS. В интактной плазмиде pAsa5 эти праймеры слишком далеко друг от друга для образования ампликона.Однако оба события гомологичной рекомбинации с участием IS могут разместить праймеры в подходящем положении для генерации ампликона. Перестройки B – C могут быть обнаружены с помощью праймеров 11B1F и 11CR, а перестройки A – C могут быть обнаружены с помощью праймеров 11AF и 11CR. Также показано положение генов traC , resD и exsD .
https://doi.org/10.1371/journal.pone.0033725.g002
Как и ожидалось, IS 11 -зависимых перегруппировок в pAsa5, которые привели к потере TTSS, были обнаружены с помощью ПЦР.На рис. 3 показано, что перегруппировки B – C и A – C были обнаружены с помощью ПЦР в производных A449. Контрольные праймеры, нацеленные на гены traC , exsD и resD (рис. 2В), были использованы для подтверждения профиля потери, ранее определенного расширенным генотипированием [10] (рис. 3). Праймеры exsD подтвердили присутствие области TTSS, тогда как праймеры traC и resD подтвердили присутствие областей выше и ниже TTSS, соответственно.Потеря сигнала ПЦР для exsD подтвердила профиль потерь типа 1 для A449-R1 (Рисунок 3A), в то время как потеря сигнала ПЦР для exsD и traC подтвердила профиль потерь типа 2 для A449-R2 (Рисунок 3B) . Перегруппировка B – C была связана с профилем потери типа 1 производного A449-R1, для которого был получен продукт ПЦР с парой праймеров 11B1F-11CR (рис. 3A). Перегруппировка A-C была связана с профилем потери типа 2 производного A449-R2, для которого был получен продукт ПЦР с парой праймеров 11AF-11CR (рис. 3B).Эти результаты показали, что перестройки, наблюдаемые в pAsa5, включают IS 11 , фланкирующие потерянную область.
Рисунок 3. IS 11. Зависимые от перегруппировки A – C и B – C участвуют в профилях потерь 2 и 1, соответственно.
Производный штамм A449-R1, несущий профиль потерь 1 ( A ), и производный штамм A449-R2, несущий профиль потерь 2 ( B ), оценивали с помощью ПЦР с использованием exsD , traC и resD . праймеры в качестве контроля (рис. 2В).Праймеры 11B1F и 11CR использовали для оценки реаранжировки B – C, в то время как праймеры 11AF и 11CR использовали для оценки реаранжировки A – C.https://doi.org/10.1371/journal.pone.0033725.g003
Мы попытались количественно оценить частоту IS-зависимых перестроек, протестировав 26 штаммов с реаранжировками pAsa5 [10], которые были получены из эталонного штамма A449 [11 ], а также два канадских изолята (01-B526 и 01-B516) с использованием праймеров, описанных выше (таблица 2). Перестройки pAsa5 с участием IS-зависимого процесса были обнаружены с использованием пар праймеров 11B1F-11CR или 11AF-11CR в девяти из 26 проанализированных производных штаммов (таблица 1).Во всех случаях полученный продукт ПЦР размером 2 т.п.н. анализировали секвенированием ампликона и выравниванием BLAST. Для каждого продукта ПЦР был идентифицирован IS 11 с фланкирующими последовательностями, подтверждающими перестройки A – C и B – C (данные не показаны).
Интересно, что все производные штаммы A449 и штаммы с профилем потери типа 2 генерировали положительный сигнал ПЦР, подтверждающий участие IS 11 в реаранжировке плазмиды pAsa5 в этих штаммах. В случае профиля потери типа 2 штамм 01-B526-R4, который потерял область TTSS, а также ген на 40 т.п.н. выше TTSS [10], претерпел реаранжировку A – C (Таблица 1).Этот необычный профиль потерь мог быть результатом неудачной попытки мобилизовать вырезанные гены после реаранжировки с участием IS 11 A и IS 11 C. Поскольку все профили потерь типа 2 были связаны с IS 11 -зависимыми перестройками, это исключает участие IS 4 , другого IS, фланкирующего область, соответствующую профилю потерь типа 2 (рис. 1).
Только шесть из 23 производных штаммов, показывающих профиль потерь типа 1, были связаны с перегруппировкой B – C посредством анализа ПЦР (Таблица 1).Профили потерь типа 1, которые нельзя объяснить перестановками B – C, могут претерпевать последующие перестройки, которые удаляют последовательности отжига 11B1F и 11CR или нарушают выравнивание праймеров 11B1F и 11CR. Это привело бы к ложноотрицательному результату ПЦР для IS 11 -зависимых перегруппировок для производных штаммов, отображающих профиль потерь типа 1, даже если перегруппировка B – C была начальным этапом процесса перегруппировки. Эти последующие непредсказуемые реаранжировки могут объяснить разницу между скоростями реаранжировки B – C и A – C, а также более высокую долю IS-зависимой реаранжировки pAsa5, идентифицированной для A449, по сравнению с канадскими изолятами.Таким образом, перегруппировки A – C могут приводить к инактивации оставшегося IS 11 в pAsa5, в то время как перегруппировки B – C могут не влиять на функцию полученного IS 11 , делая возможными последующие перегруппировки с другим IS 11 . . Это могло бы объяснить, почему профиль потери типа 1 не всегда генерировал ампликон с праймерами 11B1F и 11CR. Например, успешная перегруппировка IS с последующим другим событием, которое нарушило бы вновь образованный IS 11 B / C, могла произойти в производных 01-B526 и 01-B516, но не в штаммах производных A449, что могло бы объяснить неспособность обнаружить перестройки B – C во многих из этих штаммов.Присутствие других активных IS 11 s может привести к этим последующим перестройкам, которые нарушают выравнивание праймеров 11B1F и 11CR.
В дополнение к трем копиям IS 11 в pAsa5, копия IS 11 присутствует на хромосоме (IS 11 D), а другая — на pAsal1 (IS 11 E) [11], [ 12]. Важно отметить, что 01-B526 и 01-B516 содержат плазмиду pAsal1, а A449 — нет [11], [23]. Участие этих двух других IS 11 s, расположенных вне pAsa5, было протестировано на производных 01-B526 и 01-B516 в попытке определить, почему штаммы с профилем потерь типа 1 не генерируют ампликоны с 11B1F и Праймеры 11CR.
Для IS 11 D, который расположен на хромосоме, для ПЦР-теста использовали праймеры 11D1F2 и 11D2R2, фланкирующие IS 11 D. Если pAsa5 вставлен в хромосому через IS 11 D, расстояние между 11D1F2 и 11D2R2 должно увеличиться с 2 т.п.н. до более 117 т.п.н., предотвращая образование продукта ПЦР (рис. 4A). Встраивание pAsa5 в хромосому через IS 11 D оценивали с помощью ПЦР с использованием праймеров 11D1F2 и 11D2R2 для всех штаммов с необъяснимым профилем потери.На фигуре 4В показано, что ампликон размером 2 т.п.н. был получен во всех производных штаммах, что указывает на то, что хромосомный IS 11 D был интактным во всех штаммах и что он не участвовал в реаранжировке pAsa5.
Рисунок 4. IS 11 D на хромосоме не участвует в перестройках последовательности вставки pAsa5.
Необъяснимые перестройки в производных штаммах оценивали с помощью ПЦР с использованием праймеров 11D1F2 и 11D2R2 для областей, фланкирующих IS 11 D. A. Положения этих праймеров проиллюстрированы в обоих сценариях (интактный IS 11 D или после гомологичной рекомбинации в плазмиде pAsa5, реаранжированной B-C). B. Электрофорез продуктов ПЦР в агарозном геле, показывающий положительный результат для всех протестированных штаммов.
https://doi.org/10.1371/journal.pone.0033725.g004
Мы намеревались оценить участие IS 11 E из pAsal1 в перестройке pAsa5 путем комбинирования праймеров для областей, фланкирующих IS 11 E с праймерами 11B1F и 11CR.Положительные результаты указали бы на то, что последовательность сохранила свою непрерывность и что pAsal1 был вставлен в B – C реаранжированную pAsa5. Однако структура pAsal1 затрудняла создание специфических праймеров, поскольку pAsal1 имеет последовательность с высокой идентичностью 3,8 т.п.н. с pAsa3, другой плазмидой A. salmonicida , расположенной выше IS 11 E. Единственные существенные различия между pAsal1 и pAsa3 состоят в том, что pAsa11 содержит ген aopP TTSS и IS 11 E.Сходство между pAsal1 и pAsa3 не только усложняет дизайн праймеров для этих двух плазмид, но также может привести к трудностям секвенирования, о которых сообщалось в предыдущих исследованиях [12], [23]. Мы разработали 19 различных комбинаций праймеров для этого подхода ПЦР, ни одна из которых не была успешной, поскольку все они генерировали ампликоны ПЦР в отрицательных контролях (A449 и 01-B526). Мы также выполнили длинную ПЦР с использованием 11B1F и 11CR с праймерами для обнаружения генов выше IS 11 B и ниже IS 11 C для обнаружения возможных вставок pAsal1 в IS 11 B / C.Однако оказалось, что праймеры 11B1F и 11CR недостаточно специфичны для ПЦР с дальним действием, поскольку они генерируют большие неспецифические ампликоны в отрицательных контролях (данные не показаны). Вероятно, это связано с тем, что праймер 11B1F нацелен на область размером 5 т.п.н., которая повторяется в pAsa5, в то время как 11CR нацелена на IS 5 , который расположен непосредственно ниже IS 11 C и также многократно повторяется в A. salmonicida [11] (рисунок 1). Таким образом, было невозможно однозначно оценить рекомбинацию IS 11 E pAsal1 с IS 11 B / C pAsa5 с использованием ряда технических подходов из-за сложной и повторяющейся природы A.Структура генома salmonicida .
Необъяснимые перегруппировки в производных 01-B516 и 01-B526 могут быть результатом альтернативных IS-перегруппировок, которые также вырезают фланкирующие области. Альтернативные перегруппировки могут объяснить разницу, наблюдаемую во фракции профиля потерь типа 1, которая генерирует положительный сигнал ПЦР перегруппировки B – C с изолятами A449 и Canadian. Неправильный профиль потерь 01-B526-R4, вероятно, был вызван нерегулярной перестройкой IS. Хотя профиль потери 01-B526-R4 близко соответствовал профилю потери штамма типа 1, 01-B526-R4 также потерял часть плазмиды в 40 т.п.н. выше IS 11 B [10].Поскольку он подвергается перестройке A – C, часть вырезанной плазмиды могла быть вставлена в другую часть генома. Кроме того, перестройки B – C в канадских изолятах могли быть затронуты дефектным процессом гомологичной рекомбинации, который вырезал области, фланкирующие IS 11 , удаляя части плазмиды, которые требовались для амплификации ПЦР с использованием наших праймеров. Для Enterococcus faecalis [24] и Desulfitobacterium [25] описаны делеции смежных ДНК в результате ошибочной рекомбинации с участием IS 256 членов семейства.В обоих случаях делеции были обнаружены после воздействия на штаммы бактерий стрессовых условий.
Стрессовые условия роста оказывают пагубное влияние на целостность pAsa5 из-за присутствия в плазмиде IS 11 s. Таким образом, мы определили, подвержен ли pAsal1 стрессовым условиям роста, учитывая, что он также имеет IS 11 . Мы проверили присутствие pAsal1 в производных 01-B526 и 01-B516, используя два подхода. Сначала плазмидную ДНК из производных 01-B526 и 01-B516 очищали, расщепляли с помощью EcoRI и разделяли электрофорезом в агарозном геле.Этот метод обычно используется для обнаружения pAsal1, который мигрирует одной полосой над другими небольшими плазмидами, обычно обнаруживаемыми у A. salmonicida [12]. На рис. 5A и C показано, что четыре производных потеряли полосу pAsal1 размером 6 т.п.н. Чтобы подтвердить потерю pAsal1, тест ПЦР с использованием специфических праймеров pAsal1F и pAsal1R, нацеленных на ген aopP , специфичный для pAsal1 [12], [14] (таблица 2), показал отсутствие сигнала для тех же четырех производных штаммов (рис. 5B и D), чем те, которые демонстрируют отсутствие pAsal1 при анализе расщепления плазмиды.Это предполагает, что pAsal1 является нестабильной плазмидой, и может помочь объяснить, почему он не наблюдался во всех штаммах, оцененных другими исследователями [12], [26].
Рис. 5. Выращивание A. salmonicida в стрессовых условиях может привести к потере pAsal1.
A и C. Небольшие плазмиды (pAsa1, pAsa2, pAsa3 и pAsal1) из штаммов, полученных из 01-B526 и 01-B516, очищали и расщепляли с использованием EcoRI, что приводило к линеаризации плазмид [12]. Показан электрофорез расщепленных плазмид в агарозном геле. B и D. ДНК-лизаты тех же штаммов тестировали с помощью ПЦР с праймерами pAsal1 (таблица 2), чтобы подтвердить вторым подходом присутствие или отсутствие pAsal1 в этих штаммах. Указывается наличие (+) или отсутствие (-) продукта ПЦР.
https://doi.org/10.1371/journal.pone.0033725.g005
Заключение
В настоящем исследовании мы попытались определить, вызваны ли тепловые перестройки pAsa5, наблюдаемые у A. salmonicida , IS 11 .Мобилизация IS 11 может объяснить профили потерь типов 1 и 2 в эталонном штамме A449, которые были созданы перегруппировкой B – C и A – C, соответственно, подтверждая, что IS-зависимые перестройки происходят в A. salmonicida и что они мобильные элементы ДНК участвуют в некоторых событиях, ведущих к потере TTSS и вирулентности [10]. Наши результаты также показали, что эти перестройки могут быть вызваны стрессовыми условиями, такими как тепло, хотя это не может быть обнаружено с помощью ПЦР оптимальным образом.Таким образом, для штаммов 01-B526 и 01-B516 профили потерь не всегда можно было объяснить с помощью метода ПЦР с IS-перегруппировкой. Возможно, что гомологичная рекомбинация IS-перегруппировки в этих штаммах может быть дефектной или может включать другие события реаранжировки, что делает невозможным классифицировать их как перегруппировки B – C с использованием подхода ПЦР с IS-реаранжировкой, даже если IS 11 B и IS 11 C сыграл роль в потере области TTSS. Такие дефектные перегруппировки могли также создать уникальный профиль потерь 01-B526-R4 после события перегруппировки A – C.Дополнительная плазмида pAsal1 в канадских изолятах также могла быть вовлечена в IS-зависимые перестройки, но повторяющиеся области с высокой идентичностью, фланкирующие некоторые IS 11 s в A. salmonicida , препятствовали созданию высокоспецифичных праймеров, необходимых для ответа на этот вопрос. . Несмотря на эти ограничения, интересно отметить, что это первое исследование механистических аспектов IS-зависимых перестроек у A. salmonicida . Поскольку геном A. salmonicida содержит большое количество IS [11], будет интересно оценить пластичность A.salmonicida , который связан с IS, и определить, могут ли стрессовые условия, отличные от роста при 25 ° C, способствовать перестройке pAsa5.
Благодарности
Мы благодарим N. Turgeon (Université Laval, Quebec City, QC, Canada) за критическое прочтение рукописи. Мы благодарны C. Uhland и P. Belhumeur (Université de Montréal, Монреаль, QC, Канада) за штаммы 01-B516 и 01-B526 и M. Reith за штамм A449.
Вклад авторов
Задумал и спроектировал эксперименты: SJC KHT SDD MF.Проведены эксперименты: ХТ СДД РКД. Проанализированы данные: ХТ ГАО СДД МФ. Внесенные реактивы / материалы / инструменты анализа: ХТ СДД РКД. Написал статью: ХТ МФ ГОД.
Ссылки
- 1. Wiklund T, Dalsgaard I (1998) Возникновение и значение атипичного Aeromonas salmonicida у нелососевых и лососевых видов рыб: обзор. Dis Aquat Organ 32: 49–69.
- 2. Hiney M, Olivier G (1999) Фурункулез ( Aeromonas salmonicida ).В: Ву П, Бруно Д., редакторы. Болезни и расстройства рыб III: вирусные, бактериальные и грибковые инфекции. Оксфорд: CAB Publishing. С. 341–425.
- 3. Rasch M, Kastbjerg VG, Bruhn JB, Dalsgaard I., Givskov M, et al. (2007) Сигналы распознавания кворума вырабатываются Aeromonas salmonicida , а ингибиторы распознавания кворума могут снижать выработку потенциального фактора вирулентности. Dis Aquat Organ 78: 105–113.
- 4. Burr SE, Stuber K, Wahli T, Frey J (2002) Доказательства системы секреции типа III у Aeromonas salmonicida subsp. salmonicida . J Bacteriol 184: 5966–5970.
- 5. Альфано Дж. Р., Коллмер А. (2004) Эффекторные белки системы секреции типа III: двойные агенты при бактериальных заболеваниях и защите растений. Анну Преподобный Фитопатол 42: 385–414.
- 6. Viboud GI, Bliska JB (2005) Внешние белки Yersinia : роль в модуляции сигнальных ответов клетки-хозяина и патогенезе. Анну Рев Микробиол 59: 69–89.
- 7. Froquet R, Cherix N, Burr SE, Frey J, Vilches S и др.(2007) Альтернативная модель хозяина для оценки вирулентности Aeromonas . Appl Environ Microbiol 73: 5657–5659.
- 8. Даканай А., Никл Л., Соланки К.С., Бойд Дж. М., Уолтер Дж. А. и др. (2006) Вклад системы секреции типа III (TTSS) в вирулентность Aeromonas salmonicida subsp. salmonicida . Микробиология 152: 1847–1856.
- 9. Burr SE, Пуговкин Д., Вали Т., Сегнер Х., Фрей Дж. (2005) Ослабленная вирулентность Aeromonas salmonicida subsp.мутант по секреции типа III salmonicida в модели радужной форели. Микробиология 151: 2111–2118.
- 10. Daher RK, Filion G, Tan SG, Dallaire-Dufresne S, Paquet VE и др. (2011) Изменение факторов вирулентности и перестройка плазмиды pAsa5, вызванные ростом Aeromonas salmonicida в стрессовых условиях. Vet Microbiol 152: 353–360.
- 11. Reith ME, Singh RK, Curtis B, Boyd JM, Bouevitch A, et al. (2008) Геном Aeromonas salmonicida subsp. salmonicida A449: понимание эволюции патогена рыб. BMC Genomics 9: 427.
- 12. Бойд Дж., Уильямс Дж., Кертис Б., Козера С., Сингх Р. и др. (2003) Три маленькие криптические плазмиды из Aeromonas salmonicida subsp. salmonicida A449. Плазмида 50: 131–144.
- 13. Stuber K, Burr SE, Braun M, Wahli T, Frey J (2003) Гены секреции типа III у Aeromonas salmonicida subsp salmonicida расположены на большой термолабильной плазмиде вирулентности.J Clin Microbiol 41: 3854–3856.
- 14. Фер Д., Казанова С., Ливерман А., Блазкова Н., Орт К. и др. (2006) AopP, эффекторный белок типа III Aeromonas salmonicida , ингибирует сигнальный путь NF-kappaB. Микробиология 152: 2809–2818.
- 15. Burr SE, Stuber K, Frey J (2003) ADP-рибозилирующий токсин, AexT, из Aeromonas salmonicida subsp. salmonicida перемещается по пути секреции типа III. J Bacteriol 185: 6583–6591.
- 16. Touchon M, Rocha EP (2007) Причины обилия инсерционных последовательностей в прокариотических геномах. Mol Biol Evol 24: 969–981.
- 17. Махиллон Дж., Чендлер М. (1998) Последовательности вставки. Microbiol Mol Biol Rev 62: 725–774.
- 18. Byrne ME, Rouch DA, Skurray RA (1989) Анализ нуклеотидной последовательности IS 256 из транспозона устойчивости к гентамицин-тобрамицину-канамицину Staphylococcus aureus Tn 4001 .Gene 81: 361–367.
- 19. Шенфельдер С.М., Ланге С., Эккарт М., Хенниг С., Козицкая С. и др. (2010) Успех через разнообразие — как Staphylococcus epidermidis становится внутрибольничным патогеном. Int J Med Microbiol 300: 380–386.
- 20. Каци Е.И., Прилипов А.Г. (2009) Мобильные элементы плазмиды Azospirillum brasilense Sp245 85-MDa, участвующие в слияниях репликонов. Плазмида 62: 22–29.
- 21. Hanninen ML, Hirvela-Koski V (1997) Молекулярные и фенотипические методы для характеристики атипичных Aeromonas salmonicida .Vet Microbiol 56: 147–158.
- 22. Charette SJ, Cosson P (2004) Получение геномной ДНК из Dictyostelium discoideum для ПЦР-анализа. Биотехники 36: 574–575.
- 23. Charette SJ, Brochu F, Boyle B, Filion G, Tanaka KH и др. (2012) Проект последовательности генома вирулентного штамма 01-B526 патогена рыб Aeromonas salmonicida . J Bact 194: 722–723.
- 24. Casetta A, Hoi AB, de Cespedes G, Horaud T (1998) Разнообразие структур, несущих ген устойчивости к гентамицину высокого уровня (aac6-aph3), у штаммов Enterococcus faecalis , выделенных во Франции.Антимикробные агенты Chemother 42: 2889–2892.
- 25. Futagami T, Tsuboi Y, Suyama A, Goto M, Furukawa K (2006) Появление двух типов недехлорирующих вариантов в тетрахлорэтен-галоидыхающем Desulfitobacterium sp. штамм Y51. Appl Microbiol Biotechnol 70: 720–728.
- 26. Najimi M, Lemos ML, Osorio CR (2008) Распределение малых плазмид в Aeromonas salmonicida subsp. salmonicida штаммов, выделенных из Северо-Западного Испании и Португалии: доказательства клональности штаммов, выделенных из камбалы, Psetta maxima (L.). J Fish Dis 31: 469–472.
- 27. Dautremepuits C, Fortier M, Croisetiere S, Belhumeur P, Fournier M (2006) Модуляция клеточной иммунной системы молоди ручьей форели ( Salvelinus fontinalis ) после заражения Aeromonas salmonicida . Вет Иммунол Иммунопатол 110: 27–36.
- 28. Ebanks RO, Knickle LC, Goguen M, Boyd JM, Pinto DM, et al. (2006) Экспрессия и секреция через систему секреции Aeromonas salmonicida типа III.Микробиология 152: 1275–1286.