Какой выбрать механизм трансформации дивана
Диван является важной составляющей интерьера. Он в центре общения семьи и гостей, а в ночное время трансформируется в спальное место. Многофункциональный раскладной диван решает вопрос нехватки жилого пространства в малогабаритных квартирах. Выбор модели в таких случаях определяется не только эстетическими характеристиками, но и типом трансформации. Для ежедневного использования нужна надежная конструкция, которая выдержит частую эксплуатацию. Давайте разберемся, какие механизмы диванов бывают у производителей и какой из них лучший для сна.
Механизмы трансформации диванов
«Отелло». В основе конструкции – стальная сварная сетка и трубы. Выдерживаемая нагрузка доходит до 260 кг. Для трансформации не обязательно убирать подушки сиденья и спинки.
«Еврокнижка». Основное преимущество механизма – ровная поверхность для сна, с отсутствием перепадов. Поверхность с лицевой стороны покрыта матрасной тканью, что обеспечивает сохранение внешнего вида и долговечности обивки.
«Дельфин». Этот механизм подходит для ежедневного использования. Для трансформации нужно выдвинуть переднюю часть дивана и приподнять подъемную часть в верхнее положение.
«Миксотойл». В основе механизма – полипропиленовый тент и четыре деревянные латы. Такая конструкция подходит для нечастых раскладываний. Трансформация происходит по типу раскладушки: необходимо снять подушки с сиденья, потянуть механизм на себя и вверх, затем развернуть звенья.
«Барон». В основе механизма – фанерные площадки с настилом ППУ и металлический складной каркас. Для трансформации не требуется снимать подушки с сиденья дивана.
«Двухсекционный выкатной». В основе – усиленный каркас, выполненный из бруса хвойных пород дерева, и металлическая змейка. Она обеспечивает комфортный сон и мягкую посадку. При трансформации образуется широкое спальное место с единственным стыком.
«Еврософа». Для трансформации необходимо выдвинуть переднюю часть дивана на себя и опустить спинку в горизонтальное положение. Независимый пружинный блок позволяет получить ровную поверхность для сна с ортопедической поддержкой.
Независимый пружинный блок позволяет получить ровную поверхность для сна с ортопедической поддержкой.
«Пума». Для трансформации необходимо выдвинуть сиденье «шагающим» способом. Одновременно с этим автоматически поднимается мягкий элемент. Он находится в нише под сиденьем.
«Высоковыкатной». В основе – ортопедические латы и стальные трубы. Механизм состоит из трех сегментов, по которым равномерно распределяется нагрузка. Трансформация происходит по фрезерованным пазам в фанерном каркасе, что обеспечивает плавность и надежность работы.
«Люкс». Трансформация происходит аналогично механизму «Дельфин». Существенным отличием является ППУ, высота которого составляет 120 мм. Это соответствует высоте наполнителя основного сиденья. При трансформации получается ровное место без перепадов. Такие механизмы чаще всего используются в диванах больших габаритов. Их спальное место соответствует размерам двуспальной кровати, а иногда и превышает их.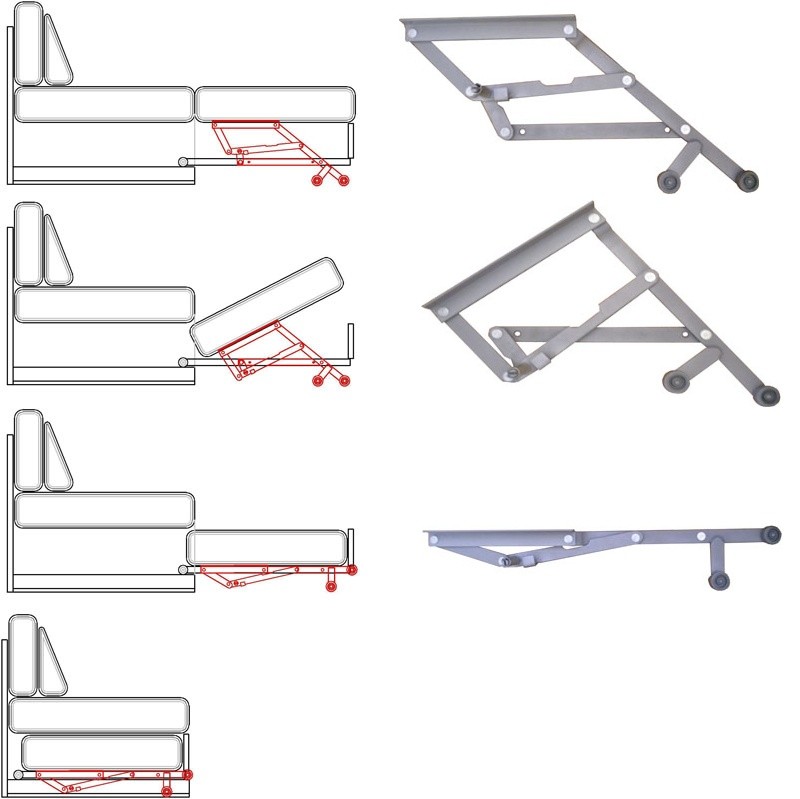
Выбор механизма раскладывания дивана
- Сопоставьте размеры мебели в сложенном и разложенном виде с площадью комнаты. Некоторые компактные конструкции в разложенном виде занимают гораздо больше места и могут загораживать выход или доступ к шкафам. Учтите размер спального места. Для комфортного сна одного человека чаще всего нужна площадь 190 см в длину и 80 см в ширину.
- Определите частоту раскладывания. Для ежедневной эксплуатации лучше выбрать максимально простой и надежный механизм, рассчитанный на большую нагрузку. Например, обратите внимание на механизм «еврокнижка», «еврософа» или «люкс», используемые компанией Home Collection при производстве диванов.
- Учтите, что некоторые механизмы раскладывания не обеспечивают ровную поверхность при трансформации.
- Подумайте, сколько человек должно удобно разместиться на диване одновременно.
 Ведь в дневное и вечернее время он часто используется как место отдыха.
Ведь в дневное и вечернее время он часто используется как место отдыха. - Решите, насколько важно для вас наличие ящиков для хранения белья. Они предусмотрены не во всех типах трансформации. Тем не менее это очень удобно в условиях малогабаритных квартир.
 Ведь в дневное и вечернее время он часто используется как место отдыха.
Ведь в дневное и вечернее время он часто используется как место отдыха.Какому бы механизму вы ни отдали предпочтение, самое важное, чтобы диван вам нравился по форме и дизайну, был удобным для отдыха и сна. Однако не стоит экономить на выборе надежного и практичного механизма, ведь мягкая мебель покупается на долгий срок. Обращайтесь за консультацией к менеджерам компании Home Collection. Они расскажут вам подробнее о каждой модели и помогут сделать выбор.
404 Not Found — Мебельная фабрика O’PRIME
Абакан Актау Алматы Анапа Архангельск Астрахань Атырау Барнаул Белгород Березники Благовещенск Бухарест Владивосток Владикавказ Владимир Волгоград Волгодонск Воронеж Геленджик Дмитров Екатеринбург Иваново Ижевск Иркутск Казань Калининград Калуга Камышин Кемерово Комсомольск-на-Амуре Костанай Кострома Краснодар Красноярск Курган Липецк Магнитогорск Майкоп Махачкала Москва Мурманск Набережные Челны Нефтеюганск Нижнекамск Нижний Новгород Новосибирск Нур-Султан Омск Орел Оренбург Пенза Пермь Петропавловск-Камчатский Прага Пятигорск Рига Ростов-на-Дону Самара Санкт-Петербург Саранск Саратов Симферополь Смоленск Сочи Ставрополь Стерлитамак Сургут Тамбов Тверь Тольятти Тула Тюмень Улан-Удэ Ульяновск Уссурийск Уфа Хабаровск Чебоксары Челябинск Череповец Чита Шахты Элиста Южно-Сахалинск Якутск Ялта Ярославль
Механизмы трансформации диванов! — Твой Дом
Раскладной диван – это не только красивый предмет мебели, но и мультифункциональное устройство, которое может решить различные задачи по компенсации недостатков помещения. При выборе важно учитывать и место, где он будет располагаться, и насколько часто будет использоваться в качестве места для сна. В зависимости от поставленных задач диван должен иметь подходящий механизм трансформации. Механизм раскладывания (или трансформации) – это своеобразные элементы, которые позволяют поменять форму и размещение мягкой мебели – преобразовать диван «для сидения» в удобное спальное место. От правильно подобранной модели зависит не только легкость и быстрота раскладывания, но и внешний вид в собранном состоянии, надежность и длительность использования.
При выборе важно учитывать и место, где он будет располагаться, и насколько часто будет использоваться в качестве места для сна. В зависимости от поставленных задач диван должен иметь подходящий механизм трансформации. Механизм раскладывания (или трансформации) – это своеобразные элементы, которые позволяют поменять форму и размещение мягкой мебели – преобразовать диван «для сидения» в удобное спальное место. От правильно подобранной модели зависит не только легкость и быстрота раскладывания, но и внешний вид в собранном состоянии, надежность и длительность использования.
1. Самым крепким и качественным является диван монолитный, в котором вообще отсутствует возможность раскладывания. Он отлично подойдет для прихожей, гостиной или маленькой кухни. Однако существенным минусом будет невозможность преобразовать диван в спальное место для двух человек.
2. «Лит» или «Эльф». Оригинальный внешне, но очень простой в использовании механизм.![]() Чтобы организовать спальное место для одного человека нужно сместить подлокотники. Отличительной особенностью также является регулировка подлокотников в подголовники. Минус модели в том, что с комфортом расположиться на ней сможет только один человек.
Чтобы организовать спальное место для одного человека нужно сместить подлокотники. Отличительной особенностью также является регулировка подлокотников в подголовники. Минус модели в том, что с комфортом расположиться на ней сможет только один человек.
3. Самым распространенным и примитивным вариантом из всех моделей является диван-книжка. Для раскладывания такой диван необходимо приподнять, услышать щелчок, затем обратно опустить. Несравненный плюс такой модели — простота раскладывания. Минус — сложенную конструкцию нужно часто двигать к стене, а при раскладывании отодвигать.
4. Более усовершенствованная модель – «клик-кляк» (также «танго»). Названа так из-за издаваемых во время трансформации звуков. От «книжки» отличается возможностью использовать не в двух, а в трех положениях – сидя, полулёжа и лёжа. Часто конструкция оснащается приподнимаемыми боковыми частями, которые используются и как подголовники, и как подлокотники.
 Минус модели в том, что сложенную конструкцию необходимо плотно придвигать к стене, как и в механизме «книжка».
Минус модели в том, что сложенную конструкцию необходимо плотно придвигать к стене, как и в механизме «книжка». 5. Еврокнижка – легко трансформируемая модель с длительным сроком службы. Большой плюс дивана — ровная поверхность для сна в разложенном виде. Механизм раскладывания очень прост – передняя часть дивана выдвигается вперед, а спинка опускается на освободившееся место сзади. Модель может быть установлена у стены, так как не требует постоянного отодвигания при раскладывании. Минусом является возможность повредить напольное покрытие при выдвигании передней части. Современные модели часто оснащены колесиками, что позволяет частично избежать этой проблемы.
6. Пантограф – усовершенствованная модель «еврокнижки». Также встречаются названия «Пума», «Тик-так», «Шагающая еврокнижка». Главное отличие от предшественника в том, что у него нет выкатных роликов. Диван раскладывается следующим образом: передняя часть поднимается и выносится вперед, после чего устанавливается на опоры. В освободившееся пространство устанавливается задняя часть. Большим плюсом является то, что диван не испортит напольное покрытие. Минус такой модели – довольно высокие цены и широкие сиденья, из-за чего невысоким людям приходится использовать дополнительные подушки.
В освободившееся пространство устанавливается задняя часть. Большим плюсом является то, что диван не испортит напольное покрытие. Минус такой модели – довольно высокие цены и широкие сиденья, из-за чего невысоким людям приходится использовать дополнительные подушки.
7. Раскладушка. Довольно сложный для раскладывания механизм. Существует несколько вариантов. Самые известные «раскладушки» — американская, французская и итальянская.
Французская (Миксотуаль, Спартак) – классическая трехсекционная модель, требующая снимания верхних подушек.
Американский вариант конструкции (Седафлекс) состоит из 2 секций и требует снимания верхних подушек для сидения.
Итальянская конструкция (также называемая «Отелло») отличается особой оригинальностью, когда трансформируется не только нижняя часть дивана, но и спинка. Механизм имеет пружинную конструкцию и усилен металлическими трубами, благодаря которым он является самым надежным из всех видов «раскладушек».
К сожалению, данные модели в целом имеют довольно много минусов. У них весьма хрупкая конструкция, в связи с чем диван может использоваться скорее как гостевой, а не постоянный. Сложная раскладка, не подходящая детям и пожилым людям. Большое количество щелей между секциями мешает комфортному отдыху. И, несмотря на довольно внушительный внешний вид, такой диван не имеет отделения для хранения белья. Плюсами таких диванов являются – просторное спальное место, удобный матрас с ортопедическим эффектом, компактность в сложенном виде.
8. Аккордеон. Модель, которая приобрела свое имя за сходство с принципом работы мехов аккордеона. Он легко раскладывается, после трансформации имеет довольно ровное и просторное спальное место. Минусом является отсутствие ящиков для хранения белья и возможность повредить напольное покрытие в случае отсутствия колес.
9. Выкатные механизмы. Такой диван – это мебельная конструкция из трёх отдельных частей. Разложить очень просто – потяните сиденье в свою сторону, благодаря чему выдвинутся все части. Затем перенесите верхнюю подушку на свободное пространство с задней стороны. Такие диваны – одни из самых надежных и долгоиграющих моделей. Минус – выкатные ролики часто повреждают пол.
Разложить очень просто – потяните сиденье в свою сторону, благодаря чему выдвинутся все части. Затем перенесите верхнюю подушку на свободное пространство с задней стороны. Такие диваны – одни из самых надежных и долгоиграющих моделей. Минус – выкатные ролики часто повреждают пол.
10. «Дельфин» (или «Кенгуру»). Одна из наиболее известных и популярных конструкций. Диваны, имеющие подобный механизм трансформации, чаще бывают угловыми, нежели прямыми. Легкие и быстрые в разборке, подходят для использования ежедневно. Практически всегда имеют ящик для хранения белья вместительного размера. Разложить диван можно в два движения. Потяните раму из-под сиденья и поднимите наверх. «Кенгуру» подобен «дельфину», с той разницей, что в разложенном виде он намного выше находится над полом. Минусом моделей является вероятность недолгой эксплуатации за счет изнашивания кронштейнов, а также часто царапающееся напольное покрытие.
11. «Конрад» («Телескоп»). Модель состоит из трех отделений. Для раскладывания потяните за нижнюю переднюю часть. Секции будут выкатываться друг за другом и встанут на специальные опоры. Механизм «Конрад» – подвид выкатного дивана и «дельфина», но отличается значительно большей высотой спального места и выкатыванием, а не выскакиванием секций. Прочная конструкция, вместительное и комфортабельное спальное место с немаленьким ящиком для хранения делают этот вид дивана одним из самых качественных вариантов. Минус – можно повредить пол при раскладывании. Также может смутить и значительный вес дивана, если его придется часто перемещать.
«Конрад» («Телескоп»). Модель состоит из трех отделений. Для раскладывания потяните за нижнюю переднюю часть. Секции будут выкатываться друг за другом и встанут на специальные опоры. Механизм «Конрад» – подвид выкатного дивана и «дельфина», но отличается значительно большей высотой спального места и выкатыванием, а не выскакиванием секций. Прочная конструкция, вместительное и комфортабельное спальное место с немаленьким ящиком для хранения делают этот вид дивана одним из самых качественных вариантов. Минус – можно повредить пол при раскладывании. Также может смутить и значительный вес дивана, если его придется часто перемещать.
12. «Реклайнер» – самая суперсовременная разновидность трансформационного механизма. В модели присутствует режим регулировки отдельных частей дивана – могут изменять свое положение спинка, подлокотники, подставка для ног, столики и т.д. В таком диване часто устанавливают дополнительные подставки, массажеры и аудиосистемы. Изготавливается такая мебель обычно из дорогих материалов и натуральной кожи, благодаря чему имеет внушительный вид. Однако устроить из такого дивана комфортное спальное место практически невозможно. Да и цена часто кусается.
Изготавливается такая мебель обычно из дорогих материалов и натуральной кожи, благодаря чему имеет внушительный вид. Однако устроить из такого дивана комфортное спальное место практически невозможно. Да и цена часто кусается.
Выбор, как вы уже заметили, огромный. У каждого механизма есть свои плюсы и минусы. Поэтому лучше ориентируйтесь на потребности Вашей семьи. Человеку, проживающему одному, будет достаточно дивана с механизмом «эльф» или «книжка». Если есть желание максимально стильно обставить дом, то можно выбрать и «реклайнер».
Семье подойдут предметы мебели, просторные в разложенном виде и аккуратные в сложенном. Также для экономии пространства стоить выбирать модели с ящиками для хранения. Не стоит выбирать диваны с ненадежными механизмами, так как они не оправдают ваших ожиданий, или неудобную модель только из-за эффектной формы и яркого цвета. Тем, кто привык спать на идеально ровной поверхности, стоит избегать конструкций, в которых образуются неровности. Также при выборе важно заранее продумать, для каких целей диван будет использоваться, где размещаться и как часто раскладываться.
Также при выборе важно заранее продумать, для каких целей диван будет использоваться, где размещаться и как часто раскладываться.
Выбрать шикарный диван
Автор статьи: Ольга Липовенко
Диван угловой Неаполь механизм Дельфин Beige 930*2450*1540(5уп)
Характеристики Описание Оплата и доставка Отзывы Наличие в магазинах
Размер (В*Ш*Г), мм: 930*2450*1540 Механизм: Дельфин Ткань: Рогожка/Искуственная кожа Бельевой ящик: Есть Спальное место: 2000*1430 Наполнитель: ППУ/Пружины змейка Наполнение подушек: Крошка ППУ, холлофайбер Материал каркаса: Фанера,ЛДСП Диван собирается на обе стороны.
-
Доставка осуществляется за порог квартиры или частного домовладения. Это подразумевает услуги грузчиков по
перемещению доставленного товара от транспорта до места удобного для осмотра товара или его временного
хранения,
включая подъем на любой этаж. Работы по демонтажу дверей и т.п. не производятся.
- В случае если товар по габаритам не проходит в дверные проёмы подъезда, квартиры или лифта, допускается его распаковка в присутствии покупателя.
- Товар доставляется как в заводской упаковке, так и в специальной транспортировочной пленке.
-
В случае невозможности доставки — либо нет подъезда на грузовой машине, качество дорожного покрытия не
соответствует безопасному проезду (провозу товара), а также невозможности осуществить доставку по другим
причинам, не зависящим от исполнителя, покупатель обязан самостоятельно забрать оплаченный товар в трехдневный
срок с момента уведомления о невозможности доставки.
 Это подразумевает услуги грузчиков по
перемещению доставленного товара от транспорта до места удобного для осмотра товара или его временного
хранения,
включая подъем на любой этаж. Работы по демонтажу дверей и т.п. не производятся.
Это подразумевает услуги грузчиков по
перемещению доставленного товара от транспорта до места удобного для осмотра товара или его временного
хранения,
включая подъем на любой этаж. Работы по демонтажу дверей и т.п. не производятся.

| г. Ростов-на-Дону, ул. 50-лет Ростсельмаша, д.1/52 | 09:00-19:00 | Под заказ | На карте |
| г. Ростов-на-Дону, ул. Красноармейская, д.157 | 10:00-22:00 | Под заказ | На карте |
| г. Ростов-на-Дону, пр. М.Нагибина, д.32Ж в ТЦ «Горизонт» | 11:00-22:00 | Под заказ | На карте |
| г. Ростов-на-Дону, ул. Малиновского, д.27а | 10:00-21:00 | Под заказ | На карте |
г. Ростов-на-Дону, пр. Космонавтов, д. 19А/28Ж в ТРЦ «ПАРК» магазин техники и мебельный салон Ростов-на-Дону, пр. Космонавтов, д. 19А/28Ж в ТРЦ «ПАРК» магазин техники и мебельный салон |
10:00-22:00 | Под заказ | На карте |
| г. Ростов-на-Дону, пр. Космонавтов, д. 19А/28Ж в ТРЦ «ПАРК» «Посудная лавка» | 10:00-22:00 | Под заказ | На карте |
| г. Ростов-на-Дону, пр. Космонавтов, д. 19А/28Ж в ТРЦ «ПАРК» магазин посуды и декора | 10:00-22:00 | Под заказ | На карте |
| г. Ростов-на-Дону, пр. Космонавтов, д. 19А/28Ж в ТРЦ «ПАРК» магазин текстиля | 10:00-22:00 | Под заказ | На карте |
Угловые диваны: выбор механизма, размеров, наполнителя
Угловые диваны появились сравнительно недавно и стали очень популярны, так что теперь и каждый вспомнит, что было время, когда о них никто и не знал. Популярность обусловлена угловым расположением данного дивана, что позволяет экономить место в квартире и использовать модель в качестве полноценной кровати.
Популярность обусловлена угловым расположением данного дивана, что позволяет экономить место в квартире и использовать модель в качестве полноценной кровати.
Угловой диван состоит из двух элементов, соединенных в форме угла. Каждая половинка представляет собой отдельно функционирующий элемент, задача которого зависит от механизма трансформации.
Про механизм
В угловом диване используются следующие механизмы (перечислены в порядке популярности):
- Еврокнижка
- Пантограф
- Дельфин или Венеция
- Аккордеон
- Седафлекс
- Трансформер
Расскажем подробней о каждом на примерах
1) Еврокнижка был и остается самым популярным механизмом как в прямых, так и в угловых моделях диванов.
Трансформация в положение кровать осуществляется выдвижением посадочного места вперед с последующим опусканием спинки на образовавшуюся часть. Просто быстро и удобно.
Просто быстро и удобно.
Важно: чтобы у выдвигающейся части ножки были на колесиках, иначе вы со временем испортите напольное покрытие.
В угловой части всегда есть ящик для белья
Также в некоторых моделях может быть ящик и в раскладывающейся части
2) Пантограф – это модифицированный вариант еврокнижки. Конструкоры доработали еврокнижку и сделали так что она больше не царапает напольное покрытие. Суть механизма его в следующем: на металлокаркас механизма прикрепляются дополнительные пружины в совокупности c механизмом. Благодаря этому посадочное место дивана не просто выдвигается вперед, а как бы шагает по дуге. Из-за этого пантограф получил альтернативное название «шагающий» или «тик-так».
Трансформация получается очень легкой, при этом посадочное место не царапает напольное покрытие.
Угловые диваны еврокнижка с пантографом стоят немного дороже, однако несомненное удобство оправдывает цену.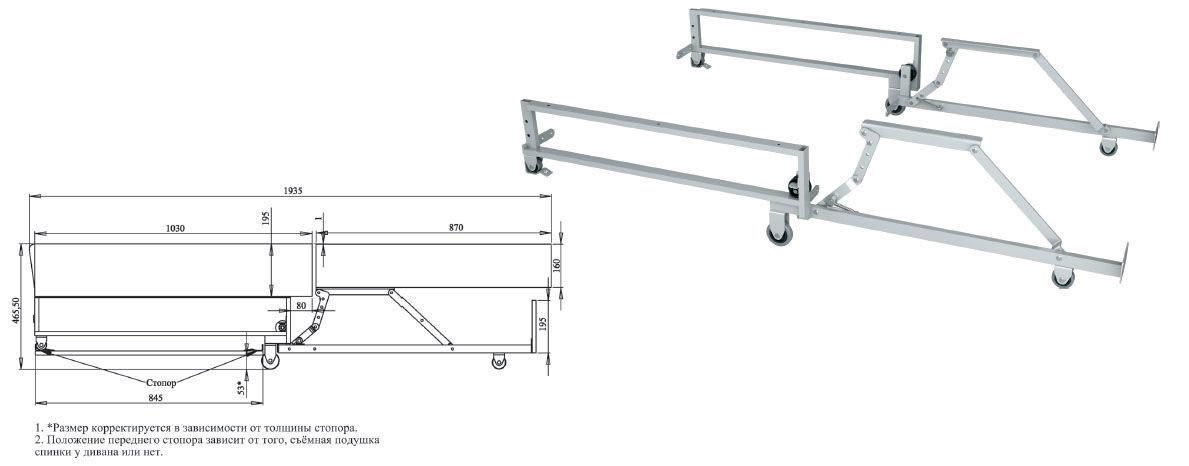
Механизмы еврокнижка и пантограф прекрасно подходят для ежедневного использования.
3) Дельфин – недавно появившийся механизм. Трансформация происходит следующим образом:
- Выдвижение короба с механизмом
- Поднятие спального места
На фото показано наглядное изображение данной трансформации.
Ящик белья в угловых диванах с механизмом дельфин есть только в угловом сегменте.
Механизм дивана дельфин прекрасно подходят для ежедневного использования.
4) Аккордеон является третьим по популярности механизмом. Раскладывается в одно движение.
Существует два существенных минуса:
1. Достаточно тяжелая трансформация ( по сути вы тяните на себя полдивана)
2. на данный каркас невозможно установить пружинный блок или ортопедический матрас.
В данной модели есть небольшой ящик под посадочным местом и в угловой части
Механизм аккордеон подходят для ежедневного использования, но трансфорация не такая легкая, как, например, в диванах еврокнижка и дельфин.
5) Седафлекс продолжает терять популярность среди механизмов, которые используются в угловых диванах. Причина: данный механизм не подходит для ежедневного раскладывания и используется исключительно как вариант гостевого дивана.
Механизм седафлекс часто называют французской раскладушкой
Ящик белья есть только в угловом сегменте.
6) Трансформер в нашем списке механизмов последний лишь по тому, что он появился совсем недавно.
Данный вариант представляет из себя разновидность прямого и углового дивана. Трансформируется все той же хорошо известной еврокнижкой.
Универсальность в товаре всегда является несомненным плюсом, поэтому диван трансформер является ХИТОМ продаж в настоящее время.
Механизм прекрасно подходит для ежедневного использования.
Полезный совет: качество дивана можно оценить по механизму. При раскладывании посадочное место не должно шататься влево-вправо. Если механизм жестко закреплен и при раскладывании ничего не шатается, значит модель качественная.
Если механизм жестко закреплен и при раскладывании ничего не шатается, значит модель качественная.
Про каркас
Каркас диванов еврокнижка, как правило из бруса, обит фанерой.
Полезный совет: качество дивана можно оценить по каркасу. Посмотрите насколько близко расположены детали дивана друг к другу. Если зазоры между подлокотником и посадочным местом минимальны, значит модель качественная. При этом обратите внимание на симметричность зазоров у двух подлокотников. Они должны быть примерно одинаковы слева и справа.
Про наполнитель
Как правило, диваны еврокнижка возможно изготовить с любым наполнителем.
Наиболее популярные:
- ППУ долговечный, но относительно жесткий. По стоимости он самый дешевый, однако, это не значит, что он менее качественный. Просто на его производство затрачивается меньше сил и материалов.

ППУ предпочитают использовать в офисах, в комнатах отдыха, в гостиницах, на кухне — везде, где имеет значение жесткость и долговечность.
- Пружинный блок с зависимыми пружинами или так называемый Боннель (Bonnel) представляет собой каркас, где в единой связи друг с другом прикреплены пружины в виде змейки.
Можно с уверенностью сказать, что за прошлое десятилетие это самый продаваемый наполнитель для дивана еврокнижка.
Данный матрас мягкий, удобный, но он менее надежный, чем ППУ. Со временем, через 7-10 лет, пружины в любом случае начнут лопаться и вылезать, прорывая обивку. При этом если лопнет одна пружина, в скором времени лопнут и близлежащие.
Как правило, данный матрас – это стандартный вариант мягкой мебели, где соотношение цена – качество – удобство — долговечность самое оптимальное.
- Ортопедический матрас или пружинный блок с независимыми пружинами представляет из себя отдельные пружины, каждая из которых в своем чехле (каркасе). Таким образом и получается независимое «поведение» данных пружин.
Данный блок очень удобный, мягкий и долговечный (срок службы до 15 лет), однако и цена у него не малая. Чем болше кличество пружин на метр квадратный, тем матрас лучше. Наиболее популярная модель 500 пружин на кв.метр.
Данный блок очень полезен для людей с заболеваниями опордо-двигательного аппрарата, а конкретно – позвоночника. Разное давление на каждую область как раз то, что нужно для больной или уставшей спины.
Полезный совет: Проверить ортопедический матрас на качество очень просто: поставьте стакан на один край дивана и сядьте неподалеку, при этом стакан не должен шелохнуться. Если у вас был такой эффект – значит перед вами настоящий ортопедический матрас
Про компоновку
Существует две существенные разновидности угловых диванов
1. Диваны с подлокотником в угловой части
2. Диваны с оттоманкой
Различие видно на фото. По сути существенной разницы между ними нет, так как набор ящиков одинаковый, и выбор зависит исключительно от ваших предпочтений.
Часто оттоманкой неверно называют вид дивана. Винтернете есть кушетка под названием «оттоманка». Это не одно и тоже.
Про ткани
Как уже было сказано ранее, Вы можете выбрать любую обивку из представленных на сайте. Различие в цене не делает ткань более качественной или менее качественной. Основная причина разницы в цене – это сложность изготовления и сложность рисунка.
Существуют следующие типы тканей: ФЛОК | ШЕНИЛЛ | ЖАККАРД | РОГОЖКА | ЭКОКОЖА | МИКРОФИБРА | МИКРОВЕЛЮР | СКОТЧГАРД
- Флок прекрасно подойдет владельцем домашних животных. Он очень плотный и надежный.
- Рогожка, или как ее иначе называют «мешковина», имеет крупное плетение и по структуре напоминает мешок, отсюда и название.
- Шенилл очень похож на тканную обивку сиденья для автомобиля. Практичен, приятен на ощупь, недорогой.
Жаккард – самая дорогая ткань. Сложный рисунок и рельефная структура делают ткань прекрасной на вид. На ощупь данный материал очень похож на шенилл. - Кожзаменитель – наиболее часто применяется для мебели на кухню или в офис. Кожзам имеет различную плотность, которая рассчитывается циклами. Если кожзам имеет более 50000 циклов, значит он вполне прослужит вам более 5-7 лет. Существуют кожзамы с 120000 циклами, например Орегон Натурал.
Таким образом, обивка это исключительно дело вкуса, стоимость которой практически не влияет на ее качество.
Что еще нужно знать.
Производство диванов еврокнижка не сложный процесс, что позволяет многим «компаниям» копировать изделия и изготавливать дешевые аналоги. Будьте внимательны и помните, что купив дешевый диван еврокнижка у недобросовестного продавца, вы можете приобрести некачественный, а главное опасный товар.
Почему опасный?
Каркас изготавливают из бруса. У больших фабрик брус приходит в закрытой машине, все детали каркаса контролируются службой ОТК.
У «копирайтеров диванов» — людей, кто копирует и ставит цены ниже, брус может быть синий и хуже того с плесенью. Выводы напрашиваются сами собой. И это только один момент, который мы здесь обсудили, касательно покупки дивана еврокнижка от производителя.
В заключение хотели бы отметить: будьте внимательны при выборе товаров.
Желаем удачного выбора и приятной покупки!
<- Назад к списку полезных статей…
ИНФОРМАЦИЯ
МЕХАНИЗМЫ
Механизм «Высоковыкатной»
Высоковыкатной, трехсекционный механизм учитывает пожелания в области создания эргономического,технологического, компактного при транспортировке и удобного при сборке дивана.
Рамка секции сиденья комплектуется би-латами, обеспечивающими наиболее комфортную посадку на диван. Остальные рамки выполнены с применением мебельной электросварной металлической сетки, обеспечивающей достаточный комфорт и надежность механизма.
Используется при изготовлении моделей: «Фернандо» прямой диван.
- Подлокотники дивана устанавливаются на основание механизма с применением быстроразъемного соединения типа «Байонет».
- Уникальная простота и легкость трансформации достигается за счет оптимального баланса кинематики механизма.
- Конструкция изголовья обеспечивает отсутствие разнотолщинности настилов в плечевой зоне в положении «кровать», что благоприятно сказывается на комфортности отдыха.
Механизм «Аккордеон ГРИД»
Слово «грид», в переводе с английского языка, означает просто-напросто сетку. Так что «Аккордеон Грид» с замком АТС – это знакомый всем механизм, но с новшествами, повышающими его потребительские свойства. В частности – с сеткой в секции сиденья.
Усовершенствование делалось с учетом сохранения внешних габаритов механизма, так что внедрение моделей на новом механизме проводится без изменения размеров и внешнего вида изделий. Это позволяет проводить быструю модернизацию модельного ряда.
Используется при изготовлении моделей с шириной механизма 80, 120, 140, 155, 180: «Дублин», «Кельн», «Паскаль», «Пегас», «Турин», «Чикаго».
- Применение мебельной металлической сетки в секции сиденья существенно улучшает эксплуатационные качества механизма и исключает поломку, даже если на сиденье дивана в сложенном состоянии встать ногами или если дети захотят превратить диван в батут.
- Усиленная конструкция рамки позволяет использовать в диване более плотные наполнители. Расположение длинных поперечных лат усиливает ортопедический эффект спального места.
- Применение металлической сетки в рамке сиденья позволило переработать способ крепления задней опоры и полностью исключить возможность её излома при складывании-раскладывании механизма, даже в случае использования в качестве наполнителя очень тяжелых материалов (например, блоки независимых пружин).
- Замок АТС Грид имеет больший угол раскрытия, нежели обычный АТС, обеспечивая еще более удобный доступ к бельевому ящику
Механизм «Аккордеон МЕККАНО»
Уникальная, запатентованная система крепления мебельной змейки к металлическому каркасу с помощью упорных змейколержателей позволила создать механизм . Теперь появилась возможность изготовить диван на механизме типа Аккордеон с необычайно комфортным местом для сидения. Используется при изготовлении моделей с шириной механизма 80, 120, 140, 155, 180 см: «Бристоль».- Механизм состоит из трех секций. Головная секция подвижно закреплена на цокольной опоре, и все три секции связаны между собой с помощью петель и уникального замка АТС.
- Обновленная конструкция замка АТС NEXT позволяет значительно увеличить угол раскрытия механизма для еще более удобного доступа к бельевому ящику и значительно снижает усилие складывания дивана.
- Применение мебельной змейки в рамке сидения позволяет теперь создать на базе механизма Аккордеон диван с уникально комфортным местом для сидения.
- Усиленная конструкция рамки позволяет использовать в диване более плотные наполнители. Расположение длинных поперечных лат усиливает ортопедический эффект спального места.
- Применение мебельной змейки в рамке сиденья позволило переработать способ крепления задней опорной ноги и полностью исключить возможность её излома при складывании-раскладывании механизма, даже в случае использования в качестве наполнителя очень тяжелых материалов (например, блоки независимых пружин).
- Применение дополнительных опорных кронштейнов позволило усилить среднюю рамку механизма в положении кровать за счет создания четырех точек опоры для нее. Данное усовершенствование, во-первых, полностью исключает возникновение скрипов в механизме во время эксплуатации дивана, во-вторых, снимает нагрузку с петель механизма, исключая вероятность их «разбалтывания», в-третьих, улучшает горизонтальность механизма в положении кровать, исключая «проседание» средней секции в процессе эксплуатации.
Механизм «Пантограф» на базе рамки «Сандра» представляет собой металлокаркасную конструкцию, состоящую из рамы сиденья и рамы спинки, изготовленных из профиля 30х20 мм с применением электросварной металлической сетки и разборного каркаса основания.
Позволяет изготовить надежный, стильный и комфортный диван-кровать, который легко трансформируется в полноценное спальное место.
Используется при изготовлении моделей: «Мадрид», «Манхэттен», «Манчестер», «Монако», «Ницца», «Пикассо», «Сиэтл».
- Все несущие рамки имеют дополнительные элементы опоры для увеличения устойчивости конструкции к деформациям при значительных нагрузках с течением времени
- Механизм предназначен для изготовления диванов с применением запатентованной безобивочной технологии, что делает чехлы таких диванов легко съемными и заменяемыми
- Конструкция рамок основания позволяет проектировать диваны с размером спального места 150х205 см (полноценная двуспальная кровать)
- В основании матраса – специализированная мебельная металлическая сетка. Обеспечивает упругость и жесткость – это дает комфорт при отдыхе и увеличивает срок службы изделия (предотвращает быструю деформацию матраса)
- Надежный механизм «Пантограф» с компенсационными пружинами делает процесс сложения/разложения легким и удобным.
- Супер легкая и быстрая сборка – подлокотники и спинка крепятся с помощью системы байонетов. Никакого инструмента не потребуется!
Механизм «Еврокнижка»
Механизм «Еврокнижка» на базе рамки «Саяна» представляет собой металлокаркасную конструкцию, состоящую из рамы сиденья и рамы спинки, изготовленных из профиля 30х20 мм с применением электросварной металлической сетки и разборного каркаса основания.
Позволяет изготовить надежный, стильный и комфортный диван-кровать, который легко трансформируется в полноценное спальное место.
Используется при изготовлении моделей: «Омега», «Виктория», «Тулуза».
- Все несущие рамки имеют дополнительные элементы опоры для увеличения устойчивости конструкции к деформациям при значительных нагрузках с течением времени
- Механизм предназначен для изготовления диванов с применением запатентованной безобивочной технологии, что делает чехлы таких диванов легко съемными и заменяемыми
- Конструкция рамок основания позволяет проектировать диваны с размером спального места 150х190 см (полноценная двуспальная кровать)
- Отсутствие эффекта опрокидывания даже при сидении на краю разложенной спинки.
- В основании матраса – специализированная мебельная металлическая сетка. Обеспечивает упругость и жесткость – это дает комфорт при отдыхе и увеличивает срок службы изделия (предотвращает быструю деформацию матраса)
- Супер легкая и быстрая сборка – подлокотники и спинка крепятся с помощью системы байонетов. Никакого инструмента не потребуется!
Механизм «Клик-Кляк» («Книжка»)
Классическая «книжка» на базе механизма «Банкетка Форте». Банкетка представляет собой хорошо отработанный, традиционный механизм типа «книжка». На сегодняшний день это один из самых популярных механизмов трансформации. Обеспечивает ровную спальную поверхность.
Используется при изготовлении моделей «Баккара», «Милан».
- Конструкция рамок основания позволяет проектировать диваны с размером спального места 140х210 см (полноценная двуспальная кровать) с очень толстым матрасом (16см)
- Расположение длинных поперечных лат усиливает ортопедический эффект спального места. Латодержатели упорного типа увеличивают срок службы матраса, создавая ровную поверхность с каркасом механизма.
- Дополнительная связь посередине способствует увеличению нагрузки на спальное место до 120 килограмм, значительно продлевая срок эксплуатации по сравнению со стандартными рамками
- Три положения для отдыха «кровать», «диван», «релакс» обеспечиваются замком NSM (замок обладает уникальными характеристиками прочности и жесткости при небольших размерах).
- Специальное инженерное решение в проектировании замка позволило добиться того, что в случае неправильной трансформации (под нагрузкой) детали NSM берут на себя основную часть нагрузки. Так что в случае поломки, потребуется заменить только замок, а не детали дивана, что существенно проще и дешевле. Заменить такой замок можно самостоятельно в домашних условиях.
Механизм «Откатной Клик-Кляк»
Ключевой особенностью этого механизма является особый замок, позволяющий раскладывать диван, не отодвигая от стены. Конструкция дает возможность использовать на спинке и сидении полноценный ортопедический матрас. Есть возможность предусмотреть вместительный ящик для белья.
Используется при изготовлении моделей «Верона».
- Конструкция рамок основания позволяет проектировать диваны с размером спального места 140х210 см (полноценная двуспальная кровать) с очень толстым матрасом (16см)
- Расположение длинных поперечных лат усиливает ортопедический эффект спального места. Латодержатели упорного типа увеличивают срок службы матраса, создавая ровную поверхность с каркасом механизма.
- Механизм комплектуется замком 3015 производства Sedac Meral Бельгия.
- Прочность конструкции значительно повышена за счет цоколя.
- Не нужно отодвигать от стены!
Механизм «Клик-Кляк» с подлокотниками-трансформерами
«Клик-кляк» на базе рамки Арджента Ретта Компатта представляет собой компактный механизм, позволяющий интегрировать в конечное изделие вместительный ящик для белья.
Механизм в положении кровать представляет собой идеально ровную спальную поверхность. Благодаря четырем складываемым независимо подлокотникам, дает возможность каждому выбрать индивидуальное положение для отдыха.
Используется при изготовлении модели «Руана».
- Конструкция рамок основания позволяет проектировать диваны с размером спального места 140х210 см (полноценная двуспальная кровать) с очень толстым матрасом (16см)
- Длинные поперечные латы усиливают ортопедический эффект спального места, увеличивают срок службы матраса, создавая ровную поверхность с каркасом механизма.
- Все подлокотники съемные, что дает высокую ремонтопригодность и меньший риск повреждения при транспортировке и хранении. Имеют 5 положений сложения.
- Три положения для отдыха «кровать», «диван», «релакс» обеспечиваются замком NSM (замок обладает уникальными характеристиками прочности и жесткости при небольших размерах).
- Специальное инженерное решение в проектировании замка позволило добиться того, что в случае неправильной трансформации (под нагрузкой) детали NSM берут на себя основную часть нагрузки. Так что в случае поломки, потребуется заменить только замок, а не детали дивана, что существенно проще и дешевле. Заменить такой замок можно самостоятельно в домашних условиях.
Механизм «Дельфин»
Цельнометаллический каркас «Просперо» в сочетании с механизмом трансформации «Дельфин» позволяют создать комфортный и удобный угловой диван с открывающейся оттоманкой, огромным ящиком для белья, выкатным механизмом спального места и стильным подлокотником. Данный механизм позволяет создавать систему модульных диванов. Конструкция механизма предусматривает смену положений оттоманки и подлокотников. Все элементы модулей соединяются между собой при помощи быстроразъемных зацепов байонет.
Используется при изготовлении угловых моделей: угловой диван «Манхэттен».
- Размер спального места 145х190 см
- Основанием спального места является мебельная сварная металлическая сетка, обеспечивающая достаточный комфорт и необыкновенную надежность механизма.
- Оттоманка с углом подъема 60 градусов обеспечивает удобный доступ к бельевому ящику.
- Металлический каркас дивана, выполненный из высокопрочных стальных труб, гарантирует соблюдение геометрии дивана на весь срок его службы и корректную и мягкую работу механизмов выдвижения и подъема.Надежные зацепы типа байонет позволяют быстро и легко соединить/разъединить элементы дивана, делая угловые модели универсальными
Механизм «Лит»
Кушетка «Лит» представляет собой компактный диванчик для студии или детской комнаты. Она отличается небольшой массой, что особенно важно в производстве мебели нового поколения. В разложенном состоянии кушетка образует идеальное ровное место, рассчитанное на одного человека.
Используется при изготовлении модели «Джимми».
- Модель «Лит» имеет подлокотники с максимальными углами подъема 90 градусов и четырьмя фиксирующимися положениями.
- Удобные подлокотники легко складываются или раскладываются, что позволяет подобрать идеальное положение индивидуально для каждого
- Высочайшая надежность металлической конструкции обеспечит долговечность и безотказность службы дивана
- Полноценное спальное место для одного человека, взрослого или ребенка
Механизм «Серджио»
Оригинальный механизм кресла-кроватей на базе металлического каркаса «Серджио». Позволяет получить кресло, которое легко и мягко трансформируется в односпальную кровать. Запатентованная разработка.
Не имеет аналогов по простоте и надежности.
Используется при изготовлении моделей: «Манхэттен», «Монако», «Ницца», «Пикассо».
- Благодаря особенностям конструкции, можно использовать целый матрас, без промежутков и швов (кроме ножной секции, где это не существенно)
- Комфортное спальное место — длина в разложенном состоянии 2 метра! (1, 97 см)
- Матрас толщиной как у дивана-кровати — 100 мм. Уникальная кинематика механизма делает процесс трансформации необыкновенно легким, а также позволяет избегать контакта элементов кресла с полом или ковролином в момент раскладывания.
- В основании матраса – специализированная мебельная металлическая сетка. Обеспечивает упругость и жесткость – это дает комфорт при отдыхе и увеличивает срок службы изделия (предотвращает быструю деформацию матраса)
- Супер легкая и быстрая сборка – подлокотники и спинка крепятся с помощью системы байонетов. Никакого инструмента не потребуется!
- На задней части каркаса предусмотрены колесики – передвинуть кресло можно быстро и без труда, исключая возможность поцарапать пол или оставить отметины на дорогом ковре.
Измерение изменяющейся во времени кинематики дельфина при всплеск-ускоренном плавании
Образец цитирования: Танака Х, Ли Дж., Утида Й, Накамура М., Икеда Т., Лю Х. (2019) Измерение изменяющейся во времени кинематики дельфина в всплеск ускоренного плавания. PLoS ONE 14 (1): e0210860. https://doi.org/10.1371/journal.pone.0210860
Редактор: Рой Гурка, Университет прибрежной Каролины, США
Поступила: 4 апреля 2018 г .; Принята к печати: 3 января 2019 г .; Опубликовано: 30 января 2019 г.
Авторские права: © 2019 Tanaka et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Все соответствующие данные находятся в документе и его файлах с вспомогательной информацией.
Финансирование: Это исследование было частично поддержано JSPS KAKENHI номер гранта JP18H05468 (HT), JP24120007 (HL), JP17K17641 (GL) и «Программой по продвижению системы отслеживания владения и пользования» Министерства образования, культуры, Спорт, наука и технологии (HT).Teral Inc. оказывала поддержку в виде заработной платы для TI, но не играла никакой дополнительной роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи. Конкретные роли этих авторов сформулированы в разделе «Авторский вклад».
Конкурирующие интересы: HL финансировалась Teral Inc. в другом совместном исследовании, не связанном с этим исследованием. TI работал на Teral Inc. HT, GL, YU и MN заявили, что никаких конкурирующих интересов не существует.Это не влияет на нашу приверженность политике PLOS ONE в отношении обмена данными и материалами.
Введение
Дельфины хорошо известны своими отличными плавательными способностями, о чем свидетельствуют высокоскоростное плавание, морские свиньи, акробатические прыжки и постановка хвоста. В частности, дельфины демонстрируют удивительно быстрое ускорение от низкой скорости до максимальной. Гидродинамика и энергетика, лежащие в основе их высокоскоростного плавания, на протяжении десятилетий привлекали широкое внимание ученых и общественности.Знаменитый «парадокс Грея» был предложен в 1936 году, в котором сэр Джеймс Грей подсчитал, что выходная мощность на килограмм мышц дельфина во время высокоскоростного плавания была в 7 раз больше, чем у человека [1]. Основываясь на этом результате, Грей также предположил, что пограничный слой вокруг дельфина может иметь ламинарный, а не турбулентный поток, что снижает сопротивление жидкости. Парадокс Грея привел к многочисленным последующим исследованиям в попытке выяснить гидродинамику плавания дельфинов [2–12]. Однако до настоящего времени не было обнаружено никаких доказательств ламинаризации пограничного слоя или другого специального механизма, снижающего сопротивление жидкости, согласно подробным обзорам Фиш и Рор (1999) [13] и Фиш (2006) [14].Фактически, расчет Грея был ошибочным из-за недооценки им мощности человека. Грей рассчитал тягу дельфина, используя наблюдаемую скорость 10,1 м / с -1 для 7-секундного спринта, тогда как мощность гребцов в течение 3-5 минут непрерывных упражнений была его эталоном для человеческих возможностей. Поскольку выходная мощность мышц со временем уменьшается, выходная мощность человека при выполнении упражнений менее 10 секунд должна быть больше, чем оценка Грея, основанная на 3–5 минутах длительных упражнений.Более того, Грей использовал в качестве показателя для сравнения мощность на единицу кг массы мышц; однако мышечную массу и механику между мышечной активностью и локомотивным движением сложно оценить.
Хотя парадокс Грея, очевидно, ложен, гидродинамический механизм движения все еще остается неясным. Одной из проблем, связанных с этим аргументом, является получение точных кинематических и гидродинамических данных для определения надежной оценки выходной мощности. Было проведено несколько предыдущих исследований, в которых сообщалось о достоверных максимальных скоростях, измеренных с использованием систематических методов.Лэнг и Дэйбелл (1963) [7] измерили максимальную скорость 7,7 м / с. -1 для Lagenorhynchus obliquidens , плавающего в прямом буксировочном резервуаре (длина 96 м, ширина 3,7 м, глубина 2,0 м). В лагуне открытого океана (размером 300 м на 35 м, глубиной 3 м) максимальная скорость 8,3 мс -1 для Tursiops truncatus gilli , плывущая на 61-метровой дистанции, сообщали Ланг и Норрис. (1966) [8]; 11,05 м с -1 для Stenella attuata , о плавании на дистанции 25 м сообщили Ланг и Прайор (1966) [15].В последнем отчете коэффициент площади сопротивления (т. Е. Сопротивление на динамическое давление) также оценивался по замедлению во время движения накатом после пересечения линии ворот. Однако результаты варьировались от 0,00288 до 0,00483 м 2 из-за неточности расчета замедления на основе данных о расстоянии и времени. Rohr et al. (2002) измерили скорость плавания различных видов дельфинов как в аквариуме, так и в дикой природе [16]. Максимальная скорость Tursiops truncatus , Delphinus delphis и Paseudorca crassidens , плавающих по периметру бассейна размером 38 м на 15 м, составила 8.2, 8.0 и 8.0 м с -1 соответственно. Максимальная скорость Tursiops truncatus непосредственно перед совершением вертикального прыжка в аквариуме составила 9,7 (стандартное отклонение) 0,8) м с -1 . Максимальная скорость свободного полета Delphinus capensis , снятая с самолета на высоте 120–145 м, составляла от 5,6 до 6,6 м с -1 .
В отличие от измерения скорости, описанного выше, изменяющееся во времени ускорение для каждого хода было плохо исследовано.Videler и Kamermans (1985) [17] зарегистрировали крейсерский полет Tursiops truncatus и Sotalia guianensis при времени менее 3,17 мс -1 в прямом резервуаре с одной кинокамерой, работающей со скоростью 50 кадров в секунду, обеспечивая последовательные боковые изображения во время двух ударов лабиринта. . Интересно, что во всех случаях дельфины ускорялись во время движений вниз, но замедлялись во время движений вверх. Совсем недавно Fish et al. (2014) [18] провели первое измерение с помощью цифровой велосиметрии с изображением частиц (DPIV), в ходе которой количественно оценили поле потока вокруг лабиринта Tursiops truncatus или позади него при старте менее 3.4 м с -1 во время нескольких ударов лабиринта. В их исследовании в качестве индикаторных частиц использовались пузырьки воздуха. Сообщалось, что каждый удар лабиринта (то есть ход вниз и ход вверх или ход вверх и ход вниз) создавал пару вращающихся в противоположных направлениях вихрей. Это означает, что гидравлическая сила была создана как при ходе вниз, так и при ходе вверх, в то время как направление силы не было определено. Однако ускорение во время каждого гребка (т. Е. Гребка вниз или вверх) при высокоскоростном плавании никогда не измерялось.Техническая задача измерения заключалась в том, чтобы зафиксировать движение плавания с достаточным разрешением для последовательных ударов лабиринта, приводящих к большому расстоянию перемещения.
В этом исследовании мы стремимся углубить понимание движительного механизма у дельфинов с помощью более точных измерений и оценок, используя современное оборудование и численный подход. В частности, мы сосредоточились на их характеристиках ускорения за один ход. Мы измерили трехмерную (трехмерную) траекторию и изменяющееся во времени ускорение дельфина Lagenorhynchus obliquidens в быстро ускоряющемся плавании в течение 7 последовательных ударов лабиринта (т.е. 14 штрихов). Дельфин, ускоряющийся в течение 2,2 с до вертикального прыжка в высоту в аквариуме, регистрировался двумя синхронизированными высокоскоростными видеокамерами, работающими со скоростью 500 кадров в секунду, что позволяло реконструировать трехмерную траекторию дельфина от начала до конца. Мы также оценили его тягу и силу тяги на основе уравнения движения и стационарной гидродинамической модели, где сопротивление дельфина рассчитывалось на основе моделирования вычислительной гидродинамики (CFD) с использованием реалистичной трехмерной жесткой модели. Чтобы получить морфологические параметры и трехмерную геометрию, еще один Lagenorhynchus obliquidens был сканирован с помощью портативного бесконтактного трехмерного сканера, и была создана точная трехмерная геометрическая модель.Коэффициенты сопротивления затем были рассчитаны консервативным способом с помощью моделирования CFD с использованием статической трехмерной модели. Комбинируя коэффициент лобового сопротивления с данными скорости и ускорения, временная тяга и мощность тяги были рассчитаны из уравнения движения. Это позволяло анализировать скорость, ускорение и тягу дельфина при каждом гребке. Результаты показали, что дельфин в основном ускорялся и создавал тягу при движении вниз, в то время как при меньшем ускорении или незначительном замедлении с меньшим усилием при подъеме вверх при всплывающем ускоренном плавании даже на высокой скорости более 5 мс -1 до более 8 мс -1 .Хотя наша оценка тяги считалась консервативной из-за постоянного моделирования CFD с жесткой моделью, результирующая максимальная усредненная по ходу тяга превышала ранее сообщаемые усредненные по времени значения в течение относительно длительного времени.
Методы
Скоростная видеозапись плывущего дельфина
Мы зафиксировали движение самки тихоокеанского белобокого дельфина ( Lagenorhynchus obliquidens ), выполняющего вертикальный прыжок в высоту в аквариуме (Yokohama Hakkeijima Sea Paradise, Yokohama, Japan) 14 декабря 2012 г.Дельфин имел длину 2,1 м и массу 110 кг. Белая мазь была нанесена на его тело в качестве маркеров для отслеживания движения (рис. 1 (A)). Примерный размер резервуара для воды составлял 35 м на 25 м, а глубина воды составляла 6 м (рис. 1 (B)). Температура воды 23 ° C. Подробная информация о благополучии животных и жилищных условиях дельфина представлена в Приложении S1.
Рис. 1. Настройка видеозаписи.
(A) Тихоокеанский белобокий дельфин ( Lagenorhynchus obliquidens ), зарегистрированный в этом исследовании.Белые мази использовались в качестве маркеров для отслеживания движения. В этом исследовании использовались маркеры клюва, боковых сторон и лабиринта, обозначенные сплошными пурпурными кружками, в то время как другие маркеры, обозначенные пунктирными голубыми кружками, не использовались. (B) Обзор резервуара для воды в Yokohama Hakkeijima Sea Paradise. Глубина воды 6 метров. На этой фотографии нет скоростных видеокамер. Вместо этого положения и направления камер указываются белыми стрелками. Плавучий остров на этой фотографии был удален, когда дельфин совершал прыжок.
https://doi.org/10.1371/journal.pone.0210860.g001
Плавание дельфина происходило следующим образом. Первоначально дельфин оставался неподвижным, высунув клюв из поверхности воды в «стартовую точку», показанную на рис. 1 (B). По сигналу дрессировщика, стоящего перед дельфином, дельфин выпрыгнул из воды в этом месте, а затем снова нырнул в воду. Дельфин спустился на дно резервуара, а затем поднялся к поверхности воды возле «точки выпрыгивания», показанной на рис. 1 (B), после чего совершил почти вертикальный прыжок.Мяч-мишень был подвешен над местом для прыжка, чтобы побудить дельфина максимально увеличить высоту прыжка. Дельфин ударил клювом по мячу на вершине прыжка. Это плавание было выполнено в рамках выставки в аквариуме. Наша запись велась во время одной из тренировок между выставками. Зарегистрированный дельфин всегда стартовал с одной и той же стартовой точки и прыгал с одной и той же точки прыжка. Высота мишени была определена аквариумом для выставки, чтобы показать самый высокий прыжок.Дельфин ежедневно обучался дотягиваться до мяча, и благодаря тренировкам высота мяча была увеличена до предела.
Обратите внимание, что мы измерили только один заплыв этого конкретного дельфина по практическим причинам, связанным с условиями аквариума. Хотя возможно, что плавательные движения, представленные в разделе «Результаты», различаются в зависимости от испытаний и отдельных лиц, из-за ежедневных повторяющихся тренировок и демонстраций плавательный режим дельфина ожидался относительно стабильным.Фактически, Rohr et al. (2002) измерили скорость плавания трех Tursiops truncatus непосредственно перед выполнением в общей сложности 47 вертикальных прыжков в аквариуме с одной подводной видеокамерой со скоростью 60 кадров в секунду, и полученная максимальная скорость варьировалась от 8,2 до 11,2 мс -1 , со средним значением 9,7 (SD 0,8) мс -1 [16].
Подводное движение дельфина было зарегистрировано двумя синхронизированными высокоскоростными видеокамерами (SA3, Photron Ltd, Япония) через прозрачную акриловую стенку резервуара для воды, как показано камерами A и B на рис. 1 (B).Третья высокоскоростная видеокамера (SA2, Photron Ltd, Япония), которая также была синхронизирована, использовалась для наблюдения за дельфином в воздухе (камера C на рис. 1 (B)). Одна из камер была выбрана в качестве главной камеры, к которой другие камеры были подключены с помощью кабелей запуска и синхронизации, обеспечивая синхронизацию для каждого кадра. Частота кадров камер была установлена на 500 кадров в секунду, а разрешение SA3 и SA2 было 1024 на 1024 пикселей и 2048 на 2048 пикселей, соответственно; кроме того, линзы для видеонаблюдения 25 мм (B2512D, Pentax Corp., Япония) использовались для всех фотоаппаратов. Примеры изображений, снятых этими тремя камерами, показаны на рис. 2. Размер правых боковых маркеров составлял около 13 см, что соответствует примерно 13 пикселям в начале погружения на захваченном изображении. Размер левого бокового маркера составлял около 5 см, что соответствует примерно 11 пикселям перед скачком на захваченном изображении. Максимальная высота левого бокового маркера оценивалась примерно в 4,5 м, принимая длину тела в качестве эталонной шкалы длины (рис. 2 (A)).
Рис. 2. Примеры изображений, снятых высокоскоростными камерами.
(A) Захваченное камерой C изображение, показывающее расчетную высоту левого бокового маркера при t = 3,248 с. (B) Наложенные изображения, снятые камерами A и B при t = 0,95 с и 2,152 с. Траектории правого и левого боковых маркеров показаны пурпурным и голубым цветом соответственно. См. Также фильмы S1 и S2.
https://doi.org/10.1371 / journal.pone.0210860.g002
Экспериментальная процедура была одобрена Комитетом по экспериментам на животных и морскому раю Йокогама Хаккейдзима Университета Тиба.
Калибровка камеры и трехмерный анализ движения
Калибровка 3-D камерыпроводилась с учетом рефракции на границе между водой и прозрачной стенкой с использованием алгоритма, предложенного Квоном [19], в котором геометрическая процедура коррекции рефракции и 2-D DLT (прямой линейный перенос ) метод были объединены.Ориентация и взаимное расположение камер и прозрачных стенок резервуара для воды были измерены с помощью лазерного дальномера (LS-511, MAX Co., Ltd., Япония). Чтобы получить восемь известных контрольных точек для калибровки стереокамеры, прямую веревку с двумя маркерами, соединяющими боковую стенку камеры и противоположную стену, сфотографировали четыре раза с изменением точек соединения.
Маркеры клюва, правой боковой, левой боковой и лапы вручную отслеживались на каждом изображении, снятом камерами, с использованием программного обеспечения для анализа движения (Dipp-Motion 3D, Ditect Co.Ltd., Япония). Исходные изображения были в 12-битной шкале серого. Вместо автоматического отслеживания было реализовано ручное отслеживание, поскольку контрастность изображений обычно не соответствовала пороговому значению программного обеспечения автоматического отслеживания даже с регулировкой контрастности и яркости изображения. Двумерные траектории боковых маркеров показаны на рис. 2 (B). Обратите внимание, что боковые маркеры и метки лапы были скрыты телом при повороте около дна. Затем были рассчитаны трехмерные координаты маркеров боковых сторон, клюва и лапы с использованием каждой пары изображений, снятых камерами.Трехмерная координата не вычислялась, если маркер был виден только в одной из камер или не виден в обеих камерах. Временные ряды полученных трехмерных координат были сглажены с использованием взвешенного скользящего среднего по 91 пункту с окном Хэмминга, предназначенного для наложения частоты среза 15 Гц для устранения шума из-за ручного отслеживания.
Поскольку центр масс дельфина был неизвестен, правый или левый боковой маркер использовался в качестве точки измерения для определения положения, скорости и ускорения дельфина в следующем анализе.Скорость и ускорение рассчитывались с использованием метода центральных разностей второго порядка следующим образом: (1) (2) где x , v x и a x — это положение, скорость и ускорение в направлении x соответственно. Верхний суффикс представляет дискретный временной индекс, а Δ t — дискретный временной интервал (0,002 с), полученный из частоты кадров высокоскоростных камер. Используя i + 2 и i-2 вместо i + 1 и i-1 в уравнениях (1) и (2), можно уменьшить шум в данных отслеживания, что приведет к более плавным данным временного ряда.
Ход вниз и вверх отличался углом отклонения тела θ , который был определен как угол между вектором маркера клюва к вектору бокового маркера и вектором вектора поперечного маркера к маркеру лабиринта в трехмерном пространстве. пробел (рис. 2 (B), внизу слева). Время начала каждого хода определялось как время, когда θ проявляли положительные пики. Визуально мы различали сгибание на вентральную сторону и сгибание на дорсальную сторону.В период, когда эти маркеры были вне поля зрения, время начала гребков определялось произвольно посредством визуального наблюдения за общим сгибанием тела.
Реконструкция реалистичной трехмерной геометрической модели дельфина (
Lagenorhynchus obliquidens )Гидродинамическое сопротивление, действующее на плавающего дельфина, было рассчитано с использованием моделирования CFD. Поскольку точное трехмерное измерение живого дельфина без какой-либо медвежьей услуги считалось невозможным, вместо этого мы измерили замороженный музейный образец того же вида ( Lagenorhynchus obliquidens ) (рис. 3).Этот экземпляр был найден мертвым на пляже залива Тояма в Японии и сохранен в холодильном хранилище Национальным музеем природы и науки Японии для инспекции. Замороженный дельфин был 1,84 м в длину и имел массу 76 кг, что соответствовало 88% и 69% от измеренного значения для живого дельфина в аквариуме, соответственно. Мы измерили трехмерную форму поверхности с помощью портативного бесконтактного трехмерного сканера (Artec Eva, Artec 3D, Люксембург) 14 апреля 2014 года. Процедура эксперимента была одобрена Национальным музеем природы и науки и Комитетом по экспериментам на животных. университета Чиба.Созданная оригинальная трехмерная модель и ее репрезентативные профили ласт, спинного плавника и ламы показаны на рис. 4 (A) и 4 (B). Поскольку форма тела была искажена при замораживании, мы реконструировали симметричную модель дельфина для моделирования CFD из исходной модели с помощью программного обеспечения 3-D CAD (Rhinoceros, Robert McNeel & Associates, США) (рис. 3 (B)) .
Рис. 4. Модели дельфинов.
(A) Оригинальная трехмерная модель замороженного дельфина. (B) Типичные формы поперечного сечения ласт, спинного плавника и лапы исходной модели. (C) Реконструированная симметричная модель, используемая для моделирования CFD. Модель была разделена на четыре части: тело, ласты, спинной плавник и плавник. См. Также модели в файлах S1 и S2.
https://doi.org/10.1371/journal.pone.0210860.g004
Процесс реконструкции симметричной модели был следующим:
- Продольная ось определялась как линия, соединяющая клюв и середину задней кромки лапы.
- Сагиттальная плоскость определялась таким образом, чтобы продольная ось лежала в сагиттальной плоскости, а спинной плавник почти совпадал со сагиттальной плоскостью.
- Исходная модель была разделена на четыре части: тело, ласты, спинной плавник и плавник.
- Построены кривые поперечного сечения тела, перпендикулярные продольной оси.
- Каждая кривая поперечного сечения тела была выровнена таким образом, чтобы дорсальный гребень поперечного сечения был выровнен в сагиттальной плоскости.
- Каждая кривая поперечного сечения была разделена сагиттальной плоскостью на левую и правую. Зеркальное отображение кривой с правой стороны было создано с левой стороны. Срединная кривая между левой кривой и зеркальной кривой была создана.
- Зеркальное отображение средней кривой, построенной на левой стороне, было создано на правой стороне. Зеркально отраженная медианная кривая на правой стороне и медианная кривая на левой стороне были объединены. Точки соединения были сглажены регулировкой локальной кривизны, и была получена симметричная кривая поперечного сечения тела.
- Симметричная поверхность тела была создана как гладкая оболочка по симметричным кривым поперечного сечения тела.
Ласты, спинной плавник и лама были отдельно реконструированы в симметричные профили. Процедура восстановления была аналогична описанной выше для тела. Для этих парных придатков в исходной модели для реконструкции использовались левый плавник и правая половина лапы, поскольку их формы считались относительно естественными.Реконструированный спинной плавник совмещен в сагиттальной плоскости. Реконструированная лапа была перпендикулярна сагиттальной плоскости и параллельна продольной оси. Реконструированные ласты были параллельны продольной оси, а угол между каждым ластом и телом был установлен на 45 градусов (рис. 4 (C)).
Морфологические параметры моделей приведены в Таблице 1 вместе с параметрами живых дельфинов, измеренными в аквариуме. Длина дельфина определялась как расстояние от клюва до середины двуустки.Реконструированные трехмерные геометрические модели дельфинов доступны в файлах S1 и S2, чтобы предоставить справочные материалы и ресурсы для будущих исследований.
Расчет коэффициента мертвого сопротивления на основе CFD
CFD-моделирование реалистичной трехмерной модели дельфина в установившемся потоке было выполнено с использованием коммерческого программного обеспечения CFD для метода конечных объемов (FVM) (ANSYS CFX R14.5, ANSYS Inc., США) для расчета сопротивления, действующего на дельфина. Основными уравнениями были трехмерное усредненное по Рейнольдсу уравнение Навье-Стокса для несжимаемой жидкости и уравнение неразрывности (т.е., сохранение массы), выражаемое как (3) (4) где i и j суффиксы обозначения тензора, u и x — средние векторы скорости и положения, ρ — плотность, p — среднее давление, а μ — вязкость. Ридебергер и Рист (2012) сообщили, что пограничный слой модели дельфина является турбулентным даже на крейсерских скоростях (3 мс -1 , Re = 1 × 10 7 ) на основе моделирования CFD с их γ — Re θ -переходная модель турбулентности [20].Чтобы обратиться к ожидаемому турбулентному пограничному слою, мы использовали модель турбулентности k — ε с масштабируемой пристеночной функцией, предоставляемой решателем. Модель турбулентности k — ε представляет собой тип модели турбулентности с усредненным по Рейнольдсу уравнением Навье-Стокса (RANS), которая широко используется для грубой оценки полей потока [21]. Ограничение модели турбулентности RANS состоит в том, что нестационарные вихри нельзя точно смоделировать и их можно недооценить [22]. Расчетный объем составлял 12 м в длину, 6 м в высоту и 3 м в ширину.Тетраэдрические сетки и 12 слоев призматических сеток, прилегающих к модели дельфина, были автоматически созданы в области жидкости с использованием программного обеспечения для построения сеток ICEM CFD, входящего в состав ANSYS (рис. 5). Область жидкости была разделена на 3 части: область с крупными ячейками, область с мелкими ячейками и область со сверхтонкими ячейками (рис. 5 (A) и 5 (B). Максимальный размер ячейки был установлен на 0,25 м для крупных ячеек). — область сетки, 0,05 м для области с мелкой сеткой и 0,01 м для области сверхтонкой сетки. Толщина первого призматического слоя, примыкающего к модели дельфина, была установлена на 0.1 мм (Рис 5 (C)). Толщина слоя увеличивалась экспоненциально в 1,2 раза, поэтому толщина самого внешнего слоя составляла 0,74 мм, а общая толщина слоев составляла 3,96 мм. Общее количество ячеек составило 7,7 × 10 6 (рис. 5 (B)). Мы использовали одни и те же сетки для всех корпусов Re . На входной границе применялась постоянная продольная скорость потока с интенсивностью турбулентности 5%. На выходной границе было установлено среднее статическое относительное давление 0 Па. Остальные стенки расчетного объема были заданы как открытые границы.Угол атаки модели дельфина, который определялся продольной осью тела, был установлен на 0 градусов во всех симуляциях.
Рис. 5. Сетки для моделирования CFD.
(A) Сетки в сагиттальной плоскости всей области жидкости. (B) Фронтальный разрез области жидкости в средней части тела. (C) Крупный план сеток в сагиттальной плоскости вблизи поверхности тела.
https://doi.org/10.1371/journal.pone.0210860.g005
Число Рейнольдса было определено как (5) где U (мс -1 ) — постоянная скорость потока, L (м) — длина дельфина, а ν (м 2 с -1 ) — кинетическая вязкость. воды при 23 ° C (9,345 × 10 −7 м 2 с -1 ). Мы выполнили CFD-моделирование с Re из 1 × 10 6 , 1 × 10 7 , 2 × 10 7 , 3 × 10 7 и 4 × 10 7 , что соответствует 0.5, 5.1, 10.2, 15.2 и 20.3 м с -1 соответственно.
Коэффициент лобового сопротивления C D рассчитывался как (6) где D — расчетное сопротивление, S поверхность — площадь поверхности дельфина, а ρ (кг · м -3 ) — плотность воды при 23 ° C (998 кг · м -3 ). Коэффициент сопротивления трения, C D , трение и коэффициент сопротивления давлением, C D , давление , были рассчитаны аналогичным образом на основе сопротивления трением и сопротивления давления, которые были отдельно выведены ANSYS. CFX.Также оценивалось сопротивление, исходящее от каждой части тела (т. Е. Тела, ласт, спинного плавника и двуустки). Обратите внимание, что коэффициент сопротивления активно плывущего дельфина ожидается выше, чем у статической модели [14]. Колеблющееся тело может уменьшить толщину пограничного слоя, что приведет к увеличению сопротивления трения до пяти раз [23]. Сопротивление давления может также увеличиваться из-за отклонения от нейтральной обтекаемой формы, когда дельфин изгибает свое тело. Более того, наша модель была полностью жесткой, в отличие от настоящего дельфина, имеющего вязкоупругую поверхность, что также может влиять на результирующее сопротивление.Следовательно, наша модель CFD предоставила консервативную оценку сопротивления, в то время как фактическое сопротивление волнообразного дельфина должно быть выше.
При использовании модели турбулентности k — ε с функцией стенки значение y + в первом призменном слое должно находиться в диапазоне логарифмического закона профиля скорости [24], который находится в диапазоне от 30 до 300 [25]. Здесь y + — это безразмерная высота от стены, выраженная как (7) где τ w (Па) — напряжение сдвига стенки, а y — высота от стены.Таким образом, y + можно определить после расчета напряжения сдвига стенки по результатам моделирования CFD. В нашем моделировании y + находились в диапазоне от 30 до 110 в большинстве мест на поверхности, когда Re было 2 × 10 7 или более, в то время как y + было ниже 30 в большинстве мест на поверхности. местоположения, когда Re было 1 × 10 7 и 1 × 10 6 (S1 Fig). Однако для слишком маленьких y + меньше 11, мы использовали масштабируемую функцию стены, которая автоматически исключила их из логарифмических вычислений.
Мы провели исследование чувствительности сетки при Re из 2 × 10 7 следующим образом. Во-первых, максимальный размер крупной ячейки 0,25 м сравнивался с размером 0,125 м. Результирующее сопротивление для случая 0,125 м отличалось от сопротивления для случая 0,25 м только на 0,2%, то есть крупная сетка 0,25 м была достаточно мелкой. Во-вторых, максимальный размер ультратонкой сетки 0,01 м сравнивался с размером 0,02 м, поскольку случай 0,005 м расходился. Результирующее сопротивление для 0.Корпус 01-м оказался меньше, чем случай 0,02-м, всего на 1,2%. Следовательно, можно сказать, что ультратонкая сетка размером 0,01 м была почти суженной. В-третьих, максимальный размер мелких ячеек был уменьшен до 0,03 м с 0,05 м, что привело к уменьшению лобового сопротивления только на 0,12%. Таким образом, было подтверждено, что мелкая ячейка 0,05 м является достаточно мелкой. Наконец, количество призматических слоев было увеличено с 12 до 17, что снизило лобовое сопротивление всего на 0,9%. Таким образом, мы использовали 12 призматических слоев.
Моделирование рывка и расчет сопротивления, тяги и мощности
Полная мгновенная сила, действующая на плывущего дельфина, была рассчитана путем умножения массы дельфина, M , на вычисленное ускорение, как описано в разделе «Методы».Составляющая тяги в направлении движения (т.е. направлении скорости), T , была рассчитана путем решения уравнений движения в направлении скорости: (8) где M — масса дельфина, — вектор ускорения, — вектор скорости, — ускорение свободного падения, V, — объем дельфина, — коэффициент сопротивления, зависящий от, — сила плавучести, и M AM — добавленная масса. Затем значение T было получено путем вычитания члена силы плавучести и силы тяжести () из суммы члена полной силы (), члена сопротивления () и члена сопротивления добавленной массы ().
M был измерен непосредственно путем взвешивания дельфина на берегу в аквариуме. S Поверхность и V дельфина в аквариуме были рассчитаны путем увеличения измерений симметричной модели CFD. Поскольку дельфин в аквариуме был в 1,14 раза больше симметричной модели по длине, масштабный коэффициент составил 1,14 2 = 1,30 для площади и 1,14 3 = 1,48 для объема. Фактически, масса в увеличенном масштабе составляла 74,4 кг × 1.48 = 110,1 кг, что почти равно измеренной массе живого дельфина в аквариуме (110 кг) (Таблица 1), что подтверждает правильность морфологического масштабирования. Таким образом, площадь поверхности и объем дельфина в аквариуме были рассчитаны и составили 1,87 м 2 и 0,111 м 3 соответственно (Таблица 1).
был рассчитан путем линейной интерполяции рассчитанных значений, полученных при моделировании CFD. Поскольку моделирование CFD было устойчивым с жесткой моделью, рассчитанное сопротивление можно рассматривать как виртуальное сопротивление паразита, которому дельфин подвергнется, если дельфин перестанет биться и перейдет в режим скольжения в статическом положении.Обратите внимание, что C D можно недооценить, как описано в разделе «Метод», что приведет к занижению при расчете T .
Добавленная масса представляет собой виртуальную массу из-за воды, окружающей дельфина. Согласно обзору Бреннена (1982), добавленная масса вытянутого сфероида в невязком потоке может быть аналитически выражена следующим образом [26]: (9) где r a — большой радиус, а r b — меньший радиус.Чтобы оценить влияние добавленной массы в нашем исследовании, мы приняли вытянутый сфероид той же длины и объема, что и наша модель дельфина (т.е. 2,1 м и 0,111 м 3 ). Затем полученные r a , r b , e , κ и M AM были 1,05 м, 0,16 м, 0,99, 0,039 и 4,3 кг соответственно. Это значение добавленной массы M AM соответствует 3,9% от M , то есть эффект добавленной массы не имеет значения в наших расчетах.
Наконец, мгновенная выходная мощность за счет тяги в направлении движения, P , была рассчитана как (10)
Обратите внимание, что расчет P не учитывает работу, выполняемую в направлении, перпендикулярном вектору скорости; таким образом, фактическая выходная мощность может быть больше P .
Расчет T и P выполнялся на каждом дискретном временном шаге с использованием значений из уравнений (1) и (2).Затем были рассчитаны усредненные значения хода и усредненные значения ударов лабиринта с использованием значений временного ряда для T и P .
Результаты
Расчет скоростей и ускорений
Для подтверждения качества калибровки камеры сравнивались исходная траектория и траектория проекции маркера клюва на плоскости изображения каждой камеры. Проекционные траектории были созданы путем проецирования рассчитанных трехмерных траекторий на плоскости изображения камеры.Как показано на рис. 6 (A) и 6 (B), траектории проекции хорошо совпадают с исходными траекториями, что указывает на достоверность калибровки камеры. Средние ошибки между исходными и проецируемыми пиксельными координатами на плоскостях изображения составляли 6,3 (SD 4,1) пикселя для камеры A и 6,4 (SD 4,0) пикселя для камеры B, соответственно.
Рис 6. Траектории маркеров.
(A, B) Проекция и исходные траектории маркера клюва для камер A и B. (C) Трехмерная траектория правого и левого строителей дельфина с числами штрихов. t = 0 с — это время, когда все тело ускользнуло под воду. Клюв достиг поверхности воды за т = 2,232 с. См. Также наборы данных S1 и S2 для трехмерных координат до и после фильтрации нижних частот.
https://doi.org/10.1371/journal.pone.0210860.g006
Полученные трехмерные траектории боковых маркеров показаны на рис. 6 (C). Ось z вертикальна, а начало координат установлено на поверхности воды. Сначала дельфин спустился и повернул направо в направлении своего движения; затем он совершил вертикальный прыжок.Как показано в дополнительных видеороликах, дельфин слегка повернулся против часовой стрелки вокруг своей продольной оси, то есть левый плавник был обращен к нижней поверхности резервуара, а дельфин повернулся на правую брюшную сторону.
Полученные угол изгиба, скорость, число Рейнольдса и ускорение показаны на рис. 7. Штрихи пронумерованы от 1 до 14, а усредненные значения хода также представлены светлыми кружками. t = 0 с соответствует моменту, когда все тело дельфина полностью проходит под водой, а t = 2.358 с соответствует времени, когда левый боковой маркер достиг поверхности воды. Первые 45 точек данных и последние 45 точек данных были обрезаны из-за 91-точечного скользящего среднего, используемого для сглаживания данных положения, как описано в разделе «Методы». Во время перехода от спуска к подъему и правый, и левый боковые маркеры были скрыты телом дельфина, поскольку он плыл к камерам. Таким образом, мы исключили этот период, от 9-го до 12-го штрихов, из последующего анализа и обсуждения.
Рис. 7. Временные характеристики измеренных параметров движения.
(A) Угол изгиба, θ . (B) Скорость и число Рейнольдса. (С) Разгон. Светлые кружки обозначают усредненные по штрихам значения. Штрихи пронумерованы от 1 до 14. Маркер лапы был вне поля зрения во время 3-го, 4-го и 9-12-го гребков. Данные о скорости и ускорении между 9-м и 12-м гребками были исключены из следующего анализа, поскольку боковые маркеры были вне поля зрения.См. Также наборы данных S3 и S4 для определения времени ключевых событий и данных временных рядов скорости и ускорения соответственно.
https://doi.org/10.1371/journal.pone.0210860.g007
Средний цикл кратковременных ударов от 1-го до 14-го гребков составлял 0,28 (стандартное отклонение 0,02) с, а средняя частота кратковременных сокращений составляла 3,7 (стандартное отклонение 0,3) Гц. Среднее отношение продолжительности хода вниз к циклу ударов лапы составило 0,49 (стандартное отклонение 0,02), что означает, что длительности ударов вниз и вверх были почти идентичны.Максимальное сгибание вентральной стороны составляло 52 ° в конце гребка №1 (ход вниз), а максимальное сгибание на тыльной стороне составляло 30 ° в конце гребка №14 (гребок вверх) (Рис. 7 (A) ).
Скорость, усредненная по ходу, быстро увеличивалась в течение первых трех ударов лапы с 5,0 мс -1 в начале 1-го гребка ( t = 0,228 с) до 8,7 мс -1 в конце 5-го гребка. ( t = 0,950 с) (Рис. 7 (B)). Максимальная средняя скорость хода составила 9.5 мс -1 при 13-м такте ( т = 1,949 с). Соответствующее усредненное по ходу число Рейнольдса варьировалось от 1,3 × 10 7 до 2,1 × 10 7 . Конечная скорость также может быть рассчитана по длине тела дельфина и продолжительности прохождения телом поверхности воды: клюв достиг поверхности воды за t = 2,232 с, задняя кромка лабиринта достигла воды за t = 2,466 с, длина тела на берегу равна 2.1 м, а затем рассчитанная скорость составила 9,0 м с -1 . Это значение составляет 95% от средней скорости 14-го гребка, равной 9,4 м / с -1 . Это различие, возможно, связано с отклонением вектора скорости левого бокового маркера от продольной оси тела, замедлением из-за потери восходящей выталкивающей силы для части тела над поверхностью воды или погрешностью измерения длины тела на берегу. Более того, ошибка в калибровке трехмерной камеры могла быть вызвана ошибкой измерения положения камеры, углов камеры или ошибкой позиционирования калибровочных маркеров из-за слабины тросов, к которым были прикреплены калибровочные маркеры.
Было обнаружено, что дельфин в основном ускорялся вперед с помощью ударов вниз, а не с помощью движений вверх, согласно усредненным значениям ускорения (рис. 7 (C)). Среднее значение среднего ускорения во время гребков вниз в первых трех ударах лавины (№ 1, 3 и 5) и стандартное отклонение составило 8,3 (SD 0,7) мс -2 (0,85 (SD 0,07) G) , тогда как во время подъема вверх в первых трех ударах лапы были -0,4 (стандартное отклонение 1,8) мс -2 (0,04 (стандартное отклонение 0,19) G).В последнем ударе лавины (№ 13 и 14) ускорение было почти нулевым как во время хода вниз, так и во время хода вверх.
CFD-потоки и коэффициенты сопротивления
Визуализация линий тока вокруг трехмерной модели дельфина при Re = 2 × 10 7 (т. Е. U = 10,2 мс -1 ) демонстрирует, что поток плавно следовал за поверхностью дельфина с этим поза во время высокоскоростного планирования (рис. 8). Скорость потока уменьшалась вокруг головы и нижней части тела и увеличивалась вокруг середины тела.Завихренность вокруг вертикальной оси и боковой оси для того же Re , показанного на фиг.9, показывает, что толщина пограничного слоя начала увеличиваться в середине тела, где площадь поперечного сечения тела максимальна. Обратите внимание, что расчет CFD установившегося потока с моделью турбулентности k — ε RANS дает грубую усредненную по времени оценку, но не может точно моделировать нестационарные вихри в пограничном слое и отрыв потока, как упоминалось в разделе «Методы».
Рис. 8. Линии тока, начинающиеся от вертикальной и горизонтальной линий в расчетах CFD с моделью белоклювого дельфина ( Re = 2 × 10 7 , U = 10,2 м с -1 ).
Цвет линий тока указывает величину скорости.
https://doi.org/10.1371/journal.pone.0210860.g008
Рис. 9. Завихренность вокруг оси y в горизонтальной плоскости (вверху) и завихренность вокруг оси z в вертикальной плоскости. (внизу) вокруг модели белоклювого дельфина ( Re = 2 × 10 7 , U = 10.2 мс -1 ).
https://doi.org/10.1371/journal.pone.0210860.g009
Расчетное давление на поверхность модели показало, что давление было положительным в голове и нижней части тела в результате уменьшения скорости потока ( Рис. 10 (A)), тогда как давление на среднюю часть корпуса было отрицательным из-за увеличения скорости потока. Обратите внимание, что 0 Па было определено как среднее статическое давление на выходной границе. Напротив, напряжение сдвига стенки было большим в верхней части корпуса и заметно уменьшилось в нижней части корпуса (рис. 10 (B)).Напряжение сдвига стенки также было высоким на каждой передней кромке киля и ласт.
Рассчитанные коэффициенты лобового сопротивления модели дельфина для различных Re сведены в Таблицу 2 вместе с соответствующими коэффициентами для каждой части. Предполагая, что твердое тело находится в установившемся потоке, было обнаружено, что как коэффициент сопротивления трения ( C D , трение ), так и коэффициент сопротивления давлением ( C D , давление ) значительно влияют на общее сопротивление. коэффициент ( C D ).Отношение сопротивления трения к общему сопротивлению составляло от 72% до 63%, а сопротивление давлению — от 28% до 37%. Также было обнаружено, что не только тело, но и другие придатки (например, ласты, спинной плавник и трематод) способствовали образованию сопротивления. Доля тела составляла от 62% до 54%, а доля других частей — от 38% до 46%.
Сила тяги и выходная мощность
Усредненная по ходу тяга в направлении движения, вычисленная по формуле (6), показана на рис. 11 (A) вместе с другими силами, такими как общая сила, сопротивление и сумма гравитационной силы и силы плавучести.Расчетная сила плавучести составляла 1075 Н, что почти компенсировало гравитационную силу 1079 Н, предполагая, что дельфин был близок к нейтральной плавучести.
Рис. 11. Расчетные силы.
(A) Значения, усредненные по ходу. (B) Средние значения Fluke-биений. Серая и белая область представляют собой ход вниз и вверх, соответственно. См. Также наборы данных S5 и S6 для получения данных о силе и коэффициенте сопротивления временного ряда и усредненных данных силы соответственно.
https://doi.org/10.1371/journal.pone.0210860.g011
Замечательная асимметрия в создании тяги между гребками вниз и вверх была обнаружена в первой половине плавания до 8-го гребка: каждый гребок вниз создавал большую тягу (от 6,7 × 10 2 до 12,5 × 10 2 Н), чем каждый ход вверх (от 0,3 × 10 2 до 3,3 × 10 2 Н). Максимальная усредненная тяга при ударе вниз соответствовала 116% веса дельфина (1079 Н).В двух последних (13-м и 14-м) тактах на максимальной скорости значения тяги (2,8 × 10 2 и 1,9 × 10 2 Н) были меньше значений сопротивления (3,6 × 10 2 и 3,6 × 10 2 Н), что приводит к отрицательным суммарным усилиям.
Тяга, усредненная по двукратным ударам, показала большие положительные значения в первых двух ударах (рис. 11 (B)). Максимальное значение составляло 7,4 × 10 2 Н, что составляет 69% от веса дельфина. Среднее сопротивление ударов стало сопоставимым с тягой при увеличении скорости, увеличиваясь с 1.5 × 10 2 до 3,6 × 10 2 N.
Мощность тяги и отношение мощности к массе показаны на рис. 12. Усредненная по ходу тяга и тяга во время хода вниз всегда были больше, чем во время хода вверх. Мощность тяги при ударах вниз варьировалась от 2,6 × 10 3 до 9,9 × 10 3 Вт, а при ударах вверх — от 0,2 × 10 3 до 2,1 × 10 3 Вт в зависимости от массы дельфина. (110 кг) отношение мощности к массе было рассчитано в диапазоне от 24 до 90 Вт кг -1 во время хода вниз и от 2 до 19 Вт кг -1 во время хода вверх.Следовательно, средняя тяговая мощность по лабиринту и среднее отношение мощности к массе составляли от 2,2 × 10 3 до 5,3 × 10 3 Вт и от 20 до 48 Вт кг -1 , соответственно.
Обсуждение
Максимальная скорость плавания
Lagenorhynchus obliquidensFish and Rohr (1999) обобщили скорость плавания различных дельфинов, включая как надежные, так и ненадежные данные; два рекорда установлены для Lagenorhynchus obliquidens по кратковременному скоростному плаванию [13].Один из них принадлежит Лэнгу и Дейбеллу (1963) [7], которые провели скоростные испытания 91-килограммового 2,0-метрового дельфина в 97-метровом резервуаре; максимальная скорость составила 7,7 м / с -1 , что составило 3,85 м / с -1 , где BL — длина его корпуса. Другой — Риджвей и Джонстон (1966) [27], которые сообщили, что дельфин может обогнать их лодку со скоростью не выше 7,2 м с -1 . Максимальная усредненная скорость хода в настоящем исследовании составила 9,5 м с -1 (4,5 BL с -1 ) на 13-м такте ( t = 1.949 с), что превышает предыдущие рекорды.
Коэффициент сопротивления
Коэффициенты сопротивления, вычисленные в настоящем исследовании, сравниваются с теоретическими значениями для плоской пластины и предыдущими данными для моделей дельфинов или дельфинов, обобщенными Fish (2014) [18] на рис. 13. Коэффициент сопротивления трением ( C D , трение ), определенный нашими расчетами, примерно соответствует теоретическому коэффициенту сопротивления трения для плоской пластины с турбулентным пограничным слоем (0.072 × Re -0,2 ). Другими словами, составляющая сопротивления давлением ( C D , давление ), по-видимому, является основным источником разницы между нашим C D и теоретическим коэффициентом сопротивления трения для плоской пластины с турбулентный пограничный слой. Значения, которые мы вычислили для C D , находятся в пределах диапазона ранее сообщенных значений для дельфинов или моделей дельфинов (черные, синие, красные и зеленые графики на рис. 13).Для более ранних CFD-симуляций дельфинов Ридебергер и Рист (2012) провели CFD-моделирование обыкновенного дельфина ( Delphinus delphis ) с использованием реалистичной трехмерной модели; вычисленное значение C D было 4 × 10 −3 для Re в диапазоне от 5,5 × 10 5 до 1 × 10 7 [20]. Наши значения C D от 6,1 × 10 −3 до 4,7 × 10 −3 для Re , равные 1 × 10 6 до 1 × 10 7 , существенно не отличаются от результаты Riedeberger et al., несмотря на то, что использовались разные виды.
Рис. 13. Расчетные коэффициенты сопротивления, C D , (белые кружки), вклад сопротивления трения в C D (незакрашенные ромбики) и вклад сопротивления давления в C D (незакрашенный квадрат) против числа Рейнольдса, наложенного на предыдущие данные, обобщенные Fish (2014) [18].Синяя линия представляет коэффициент сопротивления трения для плоской пластины с турбулентным потоком в пограничном слое, а красная линия представляет собой плоскую пластину с ламинарным пограничным потоком.Данные были получены в результате экспериментов на жестких моделях, буксируемых телах и планирующих животных (красные сплошные кружки), из гидродинамических моделей, основанных на кинематике плавания (синие квадраты), из жесткой модели «дельфин», имеющей форму твердого тела вращения. Серии NACA 66 (зеленые треугольники), а также из расчета по результатам измерений DPIV (черные сплошные круги). Данные взяты из Lang and Daybell (1963) [7]; Лэнг и Прайор (1966) [15]; Алеев и Курбатов (1974) [3]; Каян (1974) [6]; Purves et al.(1975) [10]; Уэбб (1975) [11]; Алеев (1977) [2]; Чопра и Камбэ (1977) [4]; Йейтс (1983) [12]; Виделер и Камерманс (1985) [17]; Fish (1998) [5]; и Fish et al. (2014) [18]. * Эта цифра основана на цифре Fish et al. (2014) [18].
https://doi.org/10.1371/journal.pone.0210860.g013
Наши расчеты CFD с использованием реалистичной трехмерной модели также показали, что сопротивление давления значительно влияет на общее сопротивление даже при нулевом угле атаки, как показано на Таблица 1.Обратите внимание, что ожидается, что коэффициенты сопротивления увеличиваются при увеличении угла атаки, поскольку наша модель жесткая и прямая. Более того, неустойчивое изгибание тела, как у настоящего дельфина при активном движении, может привести к увеличению коэффициентов сопротивления, как описано в разделе «Методы».
Асимметрия в ускорении и создании тяги между ходом вниз и вверх
Наиболее примечательным открытием в этом исследовании было то, что дельфин значительно ускорялся при ударах вниз, но очень мало ускорялся при ударах вверх во время кратковременного ускорения, достигая более 8 мс. -1 (удары с 1-го по 8-й на рис. 7 (C)) .Основываясь на значении C D , рассчитанном с помощью моделирования CFD, результирующие усредненные по ходу значения тяги были больше во время хода вниз, чем во время хода вверх (рис. 11 (A)). Виделер и Камерманс (1985) сообщили об аналогичной асимметрии для медленно плавающих дельфинов: их афалин ( Tursiops truncatus ) и гвианский дельфин ( Sotalia guianensis ) всегда ускорялись при спуске вниз и замедлялись при подъеме, когда плыли со скоростью 1.От 77 до 3,17 м с -1 [17]. Они также ожидали, что на высоких скоростях ход вверх может увеличить свою долю тяги и будет, по крайней мере, достаточно высоким, чтобы уравновесить сопротивление. Наш результат измерения ускорения на более высоких скоростях в диапазоне от 5,0 до 8,8 мс -1 (ходы с 1-го по 8-й), однако, не подтвердил их ожидания, но показал, что доля хода вверх в толчке все еще мала, а тяга по ходу вверх может быть меньше расчетного сопротивления.
Даже если чистая тяга (т.е. результирующая сила тяги лапы и сопротивления других частей, то есть общая сила за вычетом сил плавучести и силы тяжести в уравнении (6)) во время хода вверх была меньше, чем во время хода вниз на стадии быстрого ускорения, до 8-го хода, она все еще генерировалась значительная тяга (например, T в уравнении (6)) за исключением 6-го хода. Следовательно, принимая во внимание, что наша оценка лобового сопротивления должна быть консервативной, а волнообразный дельфин может испытывать большее сопротивление, наши наблюдения показывают, что при высокоскоростном плавании дельфин может генерировать тягу как при движении вниз, так и вверх, в то время как движение вниз, по-видимому, создает большую тягу. чем вверх.С другой стороны, во время последнего удара перед прыжком после ускорения на 9,4 мс -1 (13-й и 14-й удар), даже удар вниз не ускорял дельфина (рис. 7 (C)), что позволяет предположить, что чистая тяга Вклад движений вниз и вверх зависел как от скорости, так и от условий ускорения.
Обратите внимание, что положение маркера слежения относительно центра масс влияет на результаты скорости и ускорения. В исследовании Виделера и Кемерманса маркер располагался на спинном гребне за спинным плавником.Наш предварительный анализ, однако, показал, что значения ускорения, основанные на спинном маркере (рис. 1 (A)), были подчеркнуты, что привело к большей величине ускорения как во время хода вниз, так и вверх, чем значения, основанные на боковых маркерах. Предполагалось, что это различие связано с продольным качком и изгибом дельфина. Лэнг и Дэйбелл также сообщили, что данные временных рядов скорости в спинной части колеблются больше, чем данные в части клюва [7]. Таким образом, в настоящем исследовании были выбраны латеральные маркеры, поскольку ожидалось, что они будут ближе к центру масс на боковой проекции, чем спинной маркер.Тем не менее, наблюдалась асимметрия в генерировании тяги при ходе вниз и вверх.
Fish et al. (2014) визуализировали поле потока следа за двуусткой афалин ( Tursiops truncatus ) с помощью DPIV, используя пузырчатую пленку для медленного плавания до 3,7 м с -1 [18]. Опубликованное видео [28] показывает, что сильный вихрь выходит из лабиринта в начале хода вверх, что подразумевает создание сильной гидродинамической силы во время предыдущего хода вниз.Однако распространение вихря после подъема вверх не было подтверждено, поскольку лабиринт вышел из поля зрения видео. Вместо этого авторы отметили, что пара вращающихся в противоположных направлениях вихрей генерировалась для каждого удара лабиринта (т. Е. Хода вниз и вверх, или хода вверх и вниз) в бумаге. Их вычислительный анализ, основанный на наблюдаемой циркуляции следа, показал, что лапа генерирует гидродинамическую силу во время каждого гребка. Однако направление рассчитанной силы не уточнялось.
Таким образом, во время быстро ускоряющегося плавания в нашем наблюдении дельфин генерировал положительную чистую тягу преимущественно во время каждого хода своей лапы вниз и восстанавливался во время гребка вверх с меньшей или отрицательной чистой тягой. Оценка тяги (т.е. T в уравнении (6)) зависит от оценки сопротивления. Поскольку наша оценка C D была консервативной из-за статического моделирования CFD, расчетное усилие, следовательно, было консервативным.Для дальнейшего исследования разницы в генерировании тяги между ходом вниз и вверх желательны дальнейшие нестационарные модели CFD или гидродинамические эксперименты с движущейся моделью.
Отношение мощности к массе при быстром рывке
Рассчитанное в настоящем исследовании отношение мощности к массе сравнивалось с предыдущими данными для дельфинов и косаток при плавании со скоростью более 5 мс. -1 , как представлено в таблице 3. Таблица была построена на основе таблицы Fish и другие.(2014) [18]: случаи медленной скорости менее 5 мс -1 были удалены, добавлен случай стоячего хвоста Исогая (2014) [29], а также усреднены максимальные значения ударов и ударов. значения в настоящем исследовании были добавлены.
Размер, масса и скорость дельфина в настоящем исследовании были такими же, как в оригинальном исследовании Грэя (1936) для Delphinus delphis . Отношение мощности к массе Delphinus delphis , вычисленное Греем, предполагало, что турбулентный пограничный слой равен 21.4 Вт / кг -1 , что соответствует лишь 45% от 48 Вт / кг, определенных в нашем исследовании. Поскольку в своей оценке сопротивления жидкости Грей учитывал только сопротивление трения и пренебрежение сопротивлением давления, его оценка сопротивления жидкости и результирующей тяги занижала фактическое сопротивление на 30-40% согласно нашим расчетам CFD (Таблица 2).
Лэнг и Прайор (1966) [15] попытались измерить максимальную скорость и ускорение Stenella attuata на 25-метровом прямом курсе в лагуне размером 35 на 300 м с глубиной 4 м.Максимальная скорость была зафиксирована как 11,05 м с -1 после 2 с ускорения, а максимальная мощность наблюдалась за 0,5 с до достижения максимальной скорости. Заявленное отношение пиковой мощности к массе составило 85,7 Вт · кг -1 , что было близко к нашему максимальному усредненному значению хода 90 Вт · кг -1 , хотя в статье Лэнга и Прайора не уточняется, соответствовала ли рассчитанная мощность к усредненному значению ударов, усредненному значению хода, покадровому значению или другому показателю.
Ранее сообщаемые отношения мощности к массе для Lagenorhynchus obliquidens варьируются в зависимости от метода расчета: 67,9 Вт кг -1 с использованием квазистационарной модели Уэбба (1975) [11], 44,3 Вт кг -1 с использованием теории нестационарной подъемной поверхности Уэбба (1975) [11] и 13,4 Вт кг -1 с использованием другой теории нестационарной подъемной поверхности Йейтса (1983) [12]. Максимальное усредненное значение лавины в нашем случае (48 Вт кг -1 ) находится в пределах указанного выше диапазона, тогда как максимальное усредненное значение хода (90 Вт кг -1 ) превышает предыдущий диапазон.В общем, расчет тяги падающей лапы с использованием теории гидродинамики имеет тенденцию быть чувствительным к углу атаки лапы и характеристикам крыла (то есть кривым подъемной силы и лобового сопротивления профиля лапы). Кроме того, сложно учесть влияние нестационарных вихрей, вызванных большим углом атаки и упругой деформацией ламелей. Fish and Rohr (1999) отметили, что квазистационарная модель дает более высокие значения тяги, чем другие модели [13].Кроме того, методы, основанные на уравнениях движения тела, как в настоящем исследовании и в исследовании Лэнга и Прайора (1966), зависят от точности вычисленного ускорения и оценочного коэффициента сопротивления, используемых в уравнениях. Мгновенные значения ускорения имеют тенденцию сильно колебаться, как показано на рис. 7 (C), поскольку ускорение вычисляется как дискретный дифференциал второго порядка положения с небольшим шагом по времени. Низкочастотная фильтрация использовалась в настоящем исследовании для вычисления значений, усредненных по кратковременным биениям и по ходу, для подавления высокочастотных колебаний и получения надежных результатов.
Максимальное усредненное отношение мощности к массе 90 Вт кг -1 в настоящем исследовании является высоким по сравнению с предыдущими данными, за исключением данных Исогая (2014), который сообщил о сопоставимом значении 133,3 Вт кг — 1 for Tursiops truncatus в кратковременном стоянии на хвосте [29], при котором тело дельфина сохраняло вертикальное положение в воздухе, в то время как под водой билась только двуустка. В этом режиме плавания с опорой на хвост дельфин, как ожидается, будет генерировать большое количество энергии, чтобы выдерживать свой вес без плавучести; нет необходимости учитывать гидравлическое сопротивление, действующее на корпус, при расчете тягового усилия.Isogai создал изменяющуюся во времени трехмерную модель лапы, в которой учитывалась упругая деформация и рассчитывалась сила тяги с использованием нестационарного моделирования CFD, что позволило получить изменяющуюся во времени кривую мощности тяги. Согласно исследованию Исогая, дельфин генерировал почти симметричную положительную тягу как при движении вниз, так и вверх, а мгновенное отношение мощности к массе составляло 133,3 Вт · кг -1 , что в 2,14 раза больше, чем при ударе лабиринта. Среднее значение 62,2 Вт кг -1 .Точно так же наше максимальное усредненное значение хода 90 Вт кг -1 в 1,9 раза больше, чем максимальное усредненное значение удара лапы, равное 48 Вт кг -1 . Эти результаты предполагают, что мгновенное или усредненное отношение мощности к массе дельфинов может быть значительно больше, чем обычные усредненные значения ударов ламелей. Обратите внимание, что наша оценка C D была консервативной из-за постоянного моделирования CFD со статической моделью, поэтому наша оценка тяги и отношения мощности тяги к массе также была консервативной, а фактическая тяга и Удельная тяга может быть даже больше.В будущих исследованиях потребуется модель волнообразного тела, чтобы заменить модель твердого тела, тогда оценка лобового сопротивления, тяги и силы тяги может стать более точной, что может еще больше расширить наше понимание плавательных способностей дельфинов и механизма плавания дельфинов. .
Приложения для биомиметики
С точки зрения биомиметической инженерии, дельфины, возможно, превосходят подводные аппараты подводного типа, генерирующие тягу с обычными вращающимися гребными винтами, с точки зрения характеристик быстрого ускорения из-за их интенсивного биения лапы.При разработке биомиметических подводных роботов [31–36] не только статистически усредненные кинематические данные модельных животных, но также подробные данные временных рядов относительно конкретного человека с известной морфологией являются информативными для исследования механики и динамики передвижения. Насколько нам известно, в этой статье представлены первые кинематические данные временного ряда для конкретного дельфина с подробной морфологией в конкретном всплеск-ускоренном плавании с последовательными ударами лавины, на которые могут ссылаться разработчики биомиметических подводных роботов.Более того, расчетное отношение тяги к массе дельфина может быть использовано для сравнения характеристик тяги этих биомиметических роботов. Асимметрия в ускорении и генерировании тяги между движением вниз и вверх при плавании дельфинов особенно заслуживает дальнейшего исследования, как потенциальный источник справочной информации для разработки биомиметических плавательных роботов, приводимых в движение за счет биения хвоста.
Распознавание частотно-модулированных свистковых звуков дельфином-афалиной (Tursiops truncatus) и людьми с изменениями амплитуды, продолжительности и частоты индивидуальное признание.Здесь мы протестировали способность афалин (
Tursiops truncatus ) распознавать частотно-модулированные свистящие звуки с использованием трех альтернативных парадигм сопоставления с образцом. Дельфин сначала обучили выбирать определенный объект (объект A) в ответ на определенный звук (звук A), в общей сложности три ассоциации объект-звук. Затем звуки преобразовывались с помощью амплитуды, длительности или частотного преобразования, сохраняя при этом частотный контур каждого звука. Для сравнения, 30 участников-людей выполнили идентичное задание с одинаковыми звуками, объектами и процедурой обучения.Способность дельфина правильно сопоставлять объекты со звуками была устойчивой к изменениям амплитуды с незначительным снижением производительности в течение коротких промежутков времени. Дельфин не мог распознавать звуки, частота которых была транспонирована на плюс или минус ½ октавы. Участники продемонстрировали стойкое распознавание со всеми акустическими преобразованиями. Результаты показывают, что акустическое распознавание этим дельфином свистящих звуков ограничивалось абсолютным слухом. В отличие от человеческой речи, которая значительно различается по средней частоте, сигнатурные свистки относительно стабильны по частоте, которая могла быть выбрана для системы распознавания свистков, инвариантной к частотному транспонированию.Введение
Использование свистка дельфинами-афалинами выполняет несколько функций, включая передачу индивидуальной идентификации [1–3], голосовую метку для обращения к индивидуальным сородичам [4], поддержание групповой сплоченности [5, 6], связь на большие расстояния [7, 8] ], рекрутмент во время кормления [9] и рекламное эмоциональное состояние [10, 11]. Один тип свистка, который стал предметом многих исследований, — это «сигнатурный свисток» [2]. Подписные свистки можно определить как «усвоенный, индивидуально отличительный тип свистка в репертуаре дельфина, который раскрывает личность свиста» [12].Использование уникальной вокализации для идентификации сородичей может иметь важное значение в условиях ограниченного зрения окружающей среды океана. У дельфинов-афалин сигнатурные свистки могут составлять примерно 80–100% всех свистов, когда афалины изолированы [13], и примерно 30–70% свистов для свободно плавающих дельфинов [12]. Исследования воспроизведения показали, что дикие дельфины более склонны ориентироваться на подводного говорящего, издающего свисток более знакомого дельфина, чем на свист менее знакомого дельфина [3].Этот дифференциальный ответ часто интерпретируется как свидетельство узнавания или того, что животное научилось ассоциировать свисток со свистом. Однако можно также привести аргумент, что различное поведение дельфинов — это просто два разных ответа на знакомый и незнакомый свист без какого-либо прямого знания личности свистящего. Недвусмысленное доказательство того, что дельфины распознают ссылочный компонент сигнатурного свистка, требует прямых доказательств усвоенной связи между свистком и конкретным животным, которое издало свист.В лаборатории дельфины продемонстрировали способность понимать референтный компонент акустических (и визуальных) символов, научившись связывать определенные звуки с конкретными объектами или действиями [14, 15]. Когнитивные способности дельфинов, продемонстрированные в лаборатории, наряду с наблюдениями и экспериментами, проведенными в полевых условиях, предоставляют убедительные доказательства того, что дикие дельфины, вероятно, узнают друг друга на основе сигнатурных свистков.
Свистки дельфинов могут различаться по амплитуде и длительности, иметь повторяющиеся петли и содержать «голосовые» особенности [16].Однако стереотипный частотный контур (т. Е. График частоты свиста с течением времени), по-видимому, содержит информацию, используемую для распознавания (см.) [16]. Распознавание свиста дополнительно усложняется тем фактом, что частотный контур сигнатурного свистка может варьироваться по многим акустическим параметрам (например, количеству петель, начальной частоте, длительности, частоте точек перегиба, амплитуде и т. Д.), При этом сохраняя реляционные аспекты частотный контур [11, 17].Эти наблюдения предполагают, что способность дельфина распознавать свистки должна быть гибкой к некоторым акустическим преобразованиям. Лабораторные данные подтверждают это предположение. Например, дельфин по имени Акеакамай был обучен вокальной имитации звуков, генерируемых компьютером, с высокой степенью точности [18]. С двумя модельными звуками дельфин произвел точную имитацию, но на целую октаву выше и ниже модельного звука, что, по-видимому, является случаем октавного обобщения. В другом исследовании дельфин по имени Феникс продемонстрировал способность классифицировать последовательности тонов как имеющие восходящую или нисходящую последовательность [19].Позже она смогла обобщить эту способность на новые последовательности на полную октаву выше своих тренировочных стимулов [19]. Однако эта способность, похоже, медленно развивалась в течение четырех экспериментов после проведения тысяч испытаний. Во время первоначальных тестов передачи в эксперименте I компании Phoenix потребовались сотни испытаний в течение нескольких сеансов для достижения критериев распознавания 80% правильных ответов, и авторы пришли к выводу, что дельфин Phoenix «не обладал надежной концепцией частотного контура» на этом этапе исследование (Ralston & Herman, 1995; p271).Исследование предполагает, что распознавание частотно-сдвинутых последовательностей тонов (октавное обобщение) у дельфина Феникса не было спонтанным и явилось кульминацией длительных длительных тренировок.
Фирменный свисток дельфина СКАЗАТЬ.(A) Форма волны свистка и (B) спектрограмма того же свиста, отображающая частотный контур, который дельфины используют для распознавания. Свисток содержит несколько гармоник.
Люди легко распознают мелодии, которые были транспонированы по частоте [20], игрались на разных инструментах и игрались с разной скоростью.Дети легко учатся петь песни с помощью имитации, но часто создают песни в предпочтительном частотном диапазоне, который не соответствует точной высоте исходной модели [21]. Генерализация октав у людей хорошо известна, но редко встречается у нечеловеческих видов. Например, скворцы ( Sturnus vulgaris ), коровьи птицы ( Molothrus ater ) и пересмешники ( Mimus polyglottos ) производят сложные песни с частотной модуляцией (FM) [22]. Однако все эти птицы не смогли распознать простые частотные контуры, которые были перенесены по частоте за пределы их тренировочного диапазона [22].Система распознавания этих птиц, по-видимому, сильно ограничена абсолютным слухом [23].
Остается вопрос, насколько надежна система распознавания дельфинов для FM-свистов, которые могут изменяться по частоте, длительности и амплитуде, сохраняя при этом согласованные частотные контуры? Требует ли распознавание акустически преобразованных звуков серьезной тренировки, как у дельфинов Феникса, или их способность распознавать акустически преобразованные звуки является спонтанной? Чтобы ответить на некоторые из этих вопросов, дельфину предложили задачу с тремя вариантами сопоставления с образцом (MTS).Дельфин научился связывать три базовых FM-звука с тремя объектами, затем был протестирован со звуками, которые были преобразованы по амплитуде, длительности или частотному преобразованию, при этом сохраняя частотный контур каждого базового звука. Способность дельфина Феникса распознавать частотно-транспонированные последовательности тонов [19] предсказывает, что дельфины могут аналогичным образом распознавать частотно-транспонированные свистки. Та же самая задача MTS с использованием тех же стимулов и невербальной процедуры обучения и тестирования была представлена группе участников-людей для сравнения.Анализ ошибок между дельфином и людьми, а также постэкспериментальные интервью с людьми-участниками могут пролить свет на возможные механизмы обработки и стратегии принятия решений, которые могли использовать дельфины. Подобные сравнения человека и дельфина были сделаны в задачах распознавания эхолокации и предоставили ценную информацию об акустических сигналах, которые могли использовать дельфины [24–26].
Материалы и методы
Участники
В эксперименте участвовал атлантический афалин (SAY, женщина, 36 лет, Tursiops truncatus ) с большим опытом когнитивного [27, 28] и психофизического тестирования [29, 30].У нее была нормальная слуховая чувствительность на тестируемых частотах, как было определено с помощью тестирования слухового вызванного потенциала [31]. Ее поместили в вольеры (загоны) с плавающей сеткой размером 9 м × 9 м или 9 м × 18 м, расположенные в заливе Сан-Диего, Калифорния. Перед этим экспериментом SAY участвовал в эксперименте по маскировке слуха с использованием базовых звуков (см. Ниже), используя те же процедуры, что и в текущем эксперименте. Исследование проводилось в соответствии с протоколом, одобренным Комитетом по уходу и использованию институциональных животных Отдела биологических наук, Тихоокеанского центра космических и военно-морских боевых систем и всеми применимыми U.С. Рекомендации Министерства обороны по уходу за лабораторными животными.
Тридцать участников (20 мужчин и 10 женщин) вызвались принять участие в исследовании. Возраст участников был от 19 до 34 лет ( M = 21,8). Все участники были студентами Рочестерского технологического института или жителями Рочестера, штат Нью-Йорк. Все участники были проверены на нормальный слух с помощью теста слуха в диапазоне 250–8000 Гц (Digital Recordings, 2014). Ни один из участников не сообщил о каких-либо проблемах со слухом, и ни один из участников не показал никаких признаков потери слуха в ходе проверки слуха.Участники также заполнили анкету о своем музыкальном опыте и уровне способностей, и участники сообщили о своем музыкальном опыте от 0 до 18 лет ( M = 4,9 года) и в среднем оценили уровень способностей 2,8 по шкале от 1 до 7. Участники, завершившие исследование, получали зачет курса психологии или получали 10 долларов. Это исследование было одобрено экспертным советом Рочестерского технологического института. Перед началом эксперимента участники предоставили письменное информированное согласие.
Стимулы и процедура
Акустические стимулы дельфинов
Текущее исследование было основано на предыдущем исследовании слуховой маскировки (не описанном здесь), в котором использовались те же «базовые» FM-тоны и аналогичная экспериментальная парадигма. Те же самые FM-тоны использовались в текущем исследовании вместо реальных записанных сигнатурных свистков, чтобы избежать значительных временных затрат, связанных с переобучением дельфина новым звукам (см. Раздел «Обучение дельфинов» ниже). Три звука FM были созданы, чтобы имитировать свист дельфинов ().Один звук, обозначенный как «веревка», представлял собой линейный контур восходящей частоты. Второй звук, обозначенный как «бутылка», представлял собой повторяющуюся линейную развертку вверх, в то время как «шар» имел частотный контур, который представлял собой один цикл синусоиды. Все звуки должны быть идентичными, за исключением частотного контура, который используется для распознавания. Все базовые звуки имели длительность 500 мс с равной полосой пропускания от 8 кГц до 12 кГц с центральной частотой 10 кГц. Базовые звуки были представлены при уровне звукового давления (УЗД) 130 дБ относительно: 1 мкПа.
Базовые объекты и связанные с ними звуки.(A) «Веревка» представляла собой линейный частотный контур восходящей развертки. (B) «бутылка» представляла собой повторяющуюся линейную развертку вверх, в то время как (C) «шар» представляла собой единственный цикл синусоиды. Все базовые звуки имели длительность 500 мс с равной полосой пропускания от 8 кГц до 12 кГц с центральной частотой 10 кГц.
Тестовые звуки состояли из акустических модификаций базовых звуков в SPL, центральной частоте или продолжительности. Однако частотные контуры звуков сохранились (см.).Уровень звукового давления варьировался от 120 до 140 дБ (относительно 1 мкПа) с шагом 5 дБ, всего пять уровней. Дельфины часто издают свист с уровнем источника примерно 160 дБ относительно 1 мкПа [7, 32]. Базовый уровень 130 дБ представляет собой принимаемый уровень, который дельфин мог бы услышать, если бы он находился примерно в 30 м от объекта со свистом (при условии сферических потерь передачи). Для частотных модификаций звук был транспонирован по частоте, а не сдвинут по частоте. Частотная транспозиция является мультипликативным преобразованием и сохраняет отношение (Q) центральной частоты (CF) к ширине полосы (BW), где Q = cf / BW.Сдвиг частоты является аддитивным и сохраняет абсолютную полосу пропускания. Частотная транспозиция более уместна, чем частотный сдвиг, потому что частотное разрешение слуховой системы дельфинов имеет постоянное Q-качество на тестируемых частотах [33]. Центральные частоты были отрегулированы на ½ октавы выше и ниже базовых звуков 10 кГц, а также на 1 кГц выше и ниже базовых звуков, всего пять различных звуков. Длительность тестового звука составляла 250, 400, 500, 600 и 1000 мс, всего пять различных звуков.Изменения базовых звуков происходили только в одном акустическом измерении (например, SPL, частоте, продолжительности) за раз. Например, тестовый звук никогда не включал одновременного изменения продолжительности и SPL. Тестовые звуки состояли из 45 различных звуков ().
Примеры преобразованных тестовых звуков «мяч».(A) мяч был перемещен на ½ октавы вниз (относительно базового стимула) по частоте, (B) сдвинут на ½ октавы вверх по частоте, и (C) длительность была уменьшена вдвое до 250 мс.
Таблица 1
Тестовые звуки, использованные в эксперименте с дельфинами.Было три типа звуков (веревка, бутылка и мяч). Каждый тип звука имел три измерения звука (амплитуду, частоту и продолжительность). И каждое измерение звука имело пять различных уровней измерения, которые были протестированы, всего 45 различных звуков. Все подводные уровни дБ имели эталонный уровень 1 мкПа. Значения частоты отражают центральную частоту каждого звука.
| Размер звука | Уровень звука |
|---|---|
| амплитуда (дБ) | (дБ относительно 1 мкПа) 120, 125, 130, 135, 140 |
| частота (кГц) | (кГц ) 7.07, 9.00, 10.00, 11.00, 14.14 |
| длительность (мс) | (мс) 250, 400, 500, 600, 1000 |
Каждый звук был оцифрован, (16-битный файл Microsoft.wav с частотой дискретизации 192 кГц). Для уменьшения спектрального разбрызгивания все звуки имели линейное изменение начала-смещения 50 мс. Каждый волновой файл был преобразован в аналоговый [National Instruments USB-6251 (Austin TX)], отфильтрован [100 Гц – 100 кГц, модуль серии Krohn-Hite 3C (Броктон, Массачусетс)], ослаблен [PA5, Tucker Davis Technologies (Алачуа, Флорида) )], усиливается [Hafler P7000 (Tempe, AZ)] и проецируется в воду [пьезоэлектрический преобразователь ITC 1001 [International Transducer Corporation (Санта-Барбара, Калифорния)].Проектор ITC 1001 имел резонансную частоту 16,5 кГц с характеристикой напряжения передачи (TVR) 149 дБ относительно: 1 мкПа / В на расстоянии 1 м. Ко всем стимулам применялся корректирующий фильтр (фильтр БПФ, Cool Edit Pro 2.0) для компенсации TVR проектора и получения стимулов с плоским спектром. Генерация стимулов контролировалась с помощью специальной программы Labview. Уровни стимула были откалиброваны до 130 дБ (относительно: 1 мкПа) перед каждым сеансом путем измерения сигнала в позиции, которая будет находиться между нижней челюстью дельфина (окнами нижней челюсти) во время эксперимента.Уровни звукового давления измерялись с помощью B&K 8105 (Копенгаген, Дания, с усилением [Reson VP1000 (Slangerup, Дания)], фильтрованным [100 Гц– 100 кГц, модулем серии Krohn Hite 3C (Brockton, MA)] и оцифрованным [606,1 кГц частота обновления, разрешение 16 бит; National Instruments, USB-6251]. Сигналы снова измерялись после каждого экспериментального сеанса, и если разница между предварительной калибровкой и посткалибровкой превышала 4 дБ, данные сеанса исключались из анализа. Этого никогда не было.Когда SPL звуков был независимой переменной, SPL регулировался ручным аттенюатором [PA5, Tucker Davis Technologies (Алачуа, Флорида)].
Человеческие акустические стимулы
Участникам-людям были представлены те же базовые звуки и тестовые звуки, которые использовались с дельфином (). Однако звуки были подвергнуты пониженной дискретизации (44,1 кГц) и перенесены по частоте (с сохранением коэффициента добротности звуков) на более низкие частоты (центральная частота базовых звуков = 1000 Гц) для облегчения диапазона чувствительности человека.Регулировка амплитуды производилась путем изменения выходного напряжения с помощью Audacity 2.0.5 (Audacity, 2013). Уровни звукового давления были откалиброваны с помощью откалиброванного цифрового шумомера (Extech Instruments Model JTS-1357, 2012). Базовые звуки были представлены на уровне 70 дБ (относительно 20 мкПа), а тестовые звуки варьировались от 60 до 80 дБ с шагом 5 дБ. Вместо того, чтобы им были представлены физические объекты (мяч, бутылка, веревка), участники-люди просматривали цветные фотографии каждого объекта, напечатанные на белой бумаге (21.59 см x 27,94 см).
Таблица 2
Тестовые звуки, использованные в эксперименте на людях.Каждый тип звука имел три измерения звука (амплитуду, частоту и продолжительность). И каждое измерение звука имело пять различных уровней измерения, которые были протестированы, всего 45 различных звуков. Все уровни дБ в воздухе имели эталонный уровень 20 мкПа.
| Размер звука | Уровень звука |
|---|---|
| амплитуда (дБ) | (дБ относительно 20 мкПа) 60, 65, 70, 75, 80 |
| частота (кГц) | (Гц ) 707, 900, 1000, 1100, 1414 |
| длительность (мс) | (мс) 250, 400, 500, 600, 1000 |
Обучение дельфинов
Дельфин был обучен выполнять трехступенчатую альтернативный принудительный выбор, задача сопоставления с образцом.Во время начальной подготовки дельфин должен был стоять на подводной прикусной пластине (). Одиночный объект, нейлоновая веревка, располагался на фиксированном расстоянии 2 м от установочного устройства прикусной пластины и на той же глубине, что и прикусная пластина (примерно на 50 см ниже поверхности). Глубина укуса не была произвольной, но она давала дельфину контекстную информацию о задачах, которые она должна была выполнять. Другие накусочные пластины (например, накусочная пластина глубиной 2 м) и фиксирующие приспособления (например, накусочная пластина).g., стационарный обруч) используются для различных задач, включая задачи слухового обнаружения и задачи распознавания эхолокации. Никаких других предметов (бутылки или мяча) предъявлено не было. При предъявлении звука веревки (130 дБ) дельфин немедленно подал сигнал световым всплеском на поверхности воды (рядом с веревкой), чтобы направить внимание дельфина на нейлоновую веревку, а затем плыть и коснуться веревки своей трибуной. Когда дельфин правильно коснулся веревки в ответ на звук «веревки», в воду спроецировался звуковой сигнал «мостика», информирующий дельфина о том, что он был прав, и в этот момент он вернулся к своему дрессировщику, чтобы получить подкрепление для рыбы.После нескольких пробных тренировок сигналы splash постепенно исчезли из процедуры до тех пор, пока дельфин не стал надежно реагировать на звук веревки без сигналов. «Надежный ответ» был оперативно определен как правильный, по крайней мере, на 80% в 10 последовательных испытаниях. Затем процедура повторялась для звука бутылки и предмета из бутылки до тех пор, пока дельфин не стал надежно реагировать без подсказки. На этом этапе одновременно предъявлялись и веревка, и бутылка, и представление звука веревки или звука бутылки было рандомизировано во время каждого испытания.Веревка и бутылка всегда оставались на одних и тех же местах (). Если дельфин поплыл и коснулся нужного объекта (например, коснулся бутылки в ответ на звук бутылки), он получил перемычку зуммера, за которой следовало подкрепление рыбой. Если она коснулась не того предмета (например, коснулась веревки в ответ на звук бутылки), она получила ответный сигнал «всплеск», который указывал на неправильный выбор и велел ей вернуться на место тренера, где она не получила подкрепления рыбой. Когда она надежно реагировала и на звуки веревки, и на звуки бутылки, были введены объект мяч и звук мяча (одновременно с предыдущими объектами), и была обучена новая ассоциация звук-объект с использованием принципа исключения [34].Каждая тренировка обычно длилась от 20 до 45 минут и проводилась один или два раза в день от трех до пяти дней в неделю.
Экспериментальная ручка и предметы.(A) вид сверху и (B) вид под водой. Во время каждого испытания дельфин располагался на подводной прикусной пластине на радиальном расстоянии 2 м от нейлоновой веревки, алюминиевой бутылки, заполненной водой, и заполненной воздухом стальной сферы. Каждый из трех объектов подвешивался на моноволокне на той же глубине, что и прикусная пластина.Расположение объектов не менялось.
По завершении обучения дельфин САЙ участвовал в серии экспериментов по маскировке слуха (здесь не сообщается), в которых использовались базовые звуки и описанная выше обучаемая парадигма. Во время исследования маскировки дельфин САЙ никогда не подвергался воздействию звуков, преобразованных во времени или частоте. Однако ей были представлены базовые звуки на разных уровнях с разнообразным фоновым шумом (описание типов шума см. В Branstetter et al., 2013).В период с августа 2011 г. по апрель 2012 г. было проведено 109 дней формального обучения. В течение каждого учебного дня обычно проводилось от одного до трех занятий в зависимости от нескольких факторов, включая участие SAY в других исследовательских проектах, участие в программах повышения квалификации, оценки состояния здоровья и готовность участвовать в целом. Каждое занятие обычно состояло из 35 испытаний, но время от времени менялось по усмотрению обучающего персонала. Сбор данных из исследования слуховой маскировки проводился с мая 2012 года по сентябрь 2012 года.В период с сентября 2012 г. по январь 2013 г. проводились короткие «поддерживающие» сеансы (10–20 минут) примерно один-три раза в неделю, чтобы дельфин не забыл экспериментальную процедуру. Текущий эксперимент проводился в период с января 2013 года по март 2013 года.
Обучение людей
Процедура обучения участников-людей была разработана таким образом, чтобы отражать невербальную процедуру обучения, используемую для участников-дельфинов. Единственные устные инструкции, даваемые участникам, ориентировали их на общий объем и продолжительность задачи без указания конкретных пар объект-звук.Участники должны были найти правильные ответы путем проб и ошибок на этапе обучения, как и дельфин. Инструкции служат для ориентирования участников на основную процедуру задачи сопоставления с образцом. До настоящего исследования дельфин уже участвовал в задачах сопоставления с образцом. Точные инструкции, изложенные участникам, были следующими: «Вы будете участвовать в невербальном задании сопоставления звука и изображения. Это та же задача, которую ранее выполнял субъект-дельфин.Я могу общаться с вами только с помощью невербальных инструкций (например, жестов руками или указанием), за исключением обратной связи, которую я даю вам после того, как вы сделаете выбор. Чтобы указать свой ответ для каждого испытания, вы должны указать на картинку. Вы получите устную обратную связь (например, «правильно / неправильно») после того, как сделаете выбор. После начала сеанса я не могу отвечать ни на какие вопросы, и вам не следует использовать устное общение. Я скажу вам, когда мы пройдем половину фазы тестирования. Напоминаем, что эксперимент продлится около полутора часов.У вас есть вопросы?»
В типичном тренировочном испытании воспроизводился стимул, а затем участник касался одной из трех фотографий объекта, чтобы получить ответ. Экспериментатор дал вербальную обратную связь («правильно» или «неправильно») и сразу же приступил к следующему испытанию. В первой части тренировочного этапа перед участником размещалась только фотография с веревкой. Если участник не прикоснулся к фотографии веревки сразу после того, как услышал первый базовый звук веревки, экспериментатор либо указывал на фотографию веревки, либо брал руку участника и подносил к фотографии.Экспериментатор дал знак участнику дотронуться до веревки, указав на фотографию для трех попыток. Затем участник завершил 10 испытаний без привязки, в которых воспроизводился базовый звук веревки, и экспериментатор не указывал на фотографию веревки. Все испытания на оставшуюся часть тренировочной фазы не прошли.
На втором этапе обучения участнику была представлена фотография бутылки (фотография на веревке была удалена). Участники слышали базовый звук бутылки в десяти испытаниях и должны были указать на фотографию бутылки.Затем участнику были представлены фотографии веревки и бутылки, и они выполнили десять испытаний (по пять испытаний каждого объекта, представленных в случайном порядке). Перед тем, как перейти к следующему этапу, участники должны были набрать 90% (9 из 10 правильных ответов). Все участники набрали наивысший балл. На заключительном этапе обучения участнику были представлены все три предмета. Базовый звук мяча тренировался по принципу исключения. Заключительный этап обучения состоял из 15 испытаний (пять испытаний каждого объекта, представленных в случайном порядке), где участники должны были достичь 90% правильных ответов, чтобы перейти к этапу тестирования.Все участники соответствовали критерию ( M = 99,11%). Обучение длилось 2–4 минуты ( M = 2,93 минуты).
Процедура тестирования и анализ дельфинов
Во время каждого испытания дельфин размещался на подводной прикусной пластине () и ждал, пока будет представлен один из трех звуков (образец) (). После того, как звук был представлен, задачей дельфина было плавать и коснуться одного из трех объектов (своей трибуной), который однозначно ассоциировался со звуком (выбор). Например, если был представлен звук «веревка» (), дельфин должен был плавать и касаться веревки.Если воспроизводился звук «бутылки», дельфин должен был плавать и дотронуться до бутылки и т. Д. Тремя объектами были нейлоновая веревка (длина = 0,5 м, диаметр = 5 см), алюминиевая бутылка, наполненная водой (длина = 15 см). см, диаметр = 6 см) и стальной шар, заполненный воздухом (диаметр = 8 см). Каждый из трех объектов находился на радиальном расстоянии 2 м от прикусной пластины и подвешивался на мононити на той же глубине, что и прикусная пластина. Расположение объектов никогда не менялось, и в результате дельфин мог бы усвоить ассоциацию объекта со звуками или ассоциацию места (физическое местоположение) со звуками.Во время каждого испытания дрессировщик дельфинов выполнял роль слепого наблюдателя, поскольку она не знала о подаваемом подводном звуке, но устно сообщала ответ дельфина сотруднику-исследователю, находившемуся в соседней хижине с оборудованием. Если дельфин прикоснулся к правильному объекту, исследователь (который знал о представляемом звуке) проинструктировал дрессировщика «соединить» животное, в этот момент дельфин услышал звук зуммера (вторичное подкрепление), который заставил дельфина вернитесь на станцию дрессировщика и получите одну мойву для подкрепления.Если дельфин выбрал неправильный объект, исследователь проинструктировал дрессировщика «перезвонить» дельфину, после чего легкий всплеск руки на поверхности воды дал дельфину обратную связь о неправильном выборе и дал ей команду вернуться на станцию дрессировщика. Неправильные ответы привели к отсутствию подкрепления рыбой. Во время каждого испытания дельфин надевала на глаза присоски из латексной резины (также известные как наглазники), чтобы она не могла видеть. Таким образом, она использовала эхолокацию для навигации, идентификации и выбора каждого объекта.Основное использование наглазников состояло не в том, чтобы заставить дельфина использовать эхолокацию, а в том, чтобы помешать ей отвлекаться на зрительные отвлекающие факторы, которые, казалось, были проблемой во время фазы обучения этого эксперимента. Глазные чашки обычно используются в исследованиях эхолокации, которые показали, что дельфины могут различать и распознавать объекты и могут перемещаться по окружающей среде без использования зрения [35–37].
Каждый рабочий день проводилось от одной до трех исследовательских сессий. Каждая сессия состояла из десяти пробных разминок, пятнадцати пробных испытаний и десяти испытаний на расслабление.Разминка и заминка состояли из базовых звуков. Основная цель исходных испытаний заключалась в том, чтобы оценить мотивацию животного до и после экспериментальных испытаний, предоставив дельфину относительно «простые» испытания. Если ее производительность была менее 80% для испытаний на разминку или заминку, данные сеанса отклонялись. Испытания на заминку также помогли завершить каждую сессию на «положительной ноте», что помогло гарантировать, что дельфин будет мотивирован к участию в будущих сессиях.Базовые звуки также помогают дельфину поддерживать его работоспособность. Типы звуков (т. Е. Веревка, бутылка и мяч) были представлены в псевдослучайном порядке в любом испытании [38].
Пятнадцать тестовых испытаний состояли из тестовых звуков (), где каждый из трех типов звука был протестирован по пять раз в псевдослучайном порядке. Только одно измерение звука (то есть амплитуда, частота, продолжительность) было протестировано в течение сеанса, а пять уровней измерения были представлены в псевдослучайном порядке.Размеры звука были представлены в сбалансированном формате ABBA (то есть амплитуда, частота, продолжительность, продолжительность, частота и амплитуда), чтобы уменьшить любые потенциальные эффекты обучения. Каждый из 45 тестовых звуков был протестирован по десять раз каждый, в общей сложности 450 тестовых попыток.
Статистический анализ проводился с использованием R [39]. Полиномиальная регрессия использовалась, чтобы определить, правильно ли соотносятся акустический размер и тип звука с пропорцией. Вместо линейных моделей были выбраны полиномиальные модели, потому что тестовые звуки как увеличивались, так и уменьшались (в SPL, частоте или продолжительности) от базовых звуков, что приводило к большим различиям ПК по мере того, как уровни акустических размеров отклонялись от базовых значений.Это привело к правильным функциям перевернутой U-образной пропорции. Упрощение модели было выполнено с использованием пошагового удаления, чтобы определить, какие переменные-предикторы были значимыми.
Процедура тестирования и анализ на людях
Один экспериментатор тестировал участников в тихой комнате. Стимулы предъявлялись участнику и экспериментатору через два набора накладных наушников Bose (Bose Corp, 2009) с портативного компьютера MacBook Pro (Apple Inc., 2011). Звуки воспроизводились с помощью QuickTime Player версии 7.6.6 (Apple Inc., 2010). Участник сидел лицом к экспериментатору и фотографиям, но не мог видеть экран компьютера. Перед началом процедуры обучения участников проинструктировали, что они будут выполнять невербальную задачу сопоставления звука и изображения (см. Выше). Никаких дальнейших устных инструкций до начала фазы тестирования не давалось. Задача была разработана так, чтобы быть как можно более похожей на трехальтернативную задачу сопоставления с образцом с принудительным выбором, выполняемую субъектом-дельфином, за исключением того, что участники завершили интервью после завершения этапов обучения и тестирования.
Этап тестирования начался сразу после завершения обучения. Всего было 18 тестовых сессий (6 подходов по 3 занятия в каждом). Каждая серия из 3 сеансов содержала только один тип акустической трансформации (амплитуда, продолжительность или частота). Порядок проведения 18 сессий соответствовал уравновешенному дизайну ABBA. Участников случайным образом распределили по одному из трех порядков. Порядок А соответствовал порядку сеансов, используемому для субъекта-дельфина (набор амплитуд, набор частот, два набора продолжительности, набор частот, набор амплитуд).Порядок B начинается и заканчивается набором длительности (набор длительности, набор амплитуды, два набора частот, набор амплитуд, набор длительности), а порядок C начинается и заканчивается набором частоты (набор частоты, набор длительности, два набора амплитуд, набор длительности, набор частот).
Состав сеанса был точно таким же для дельфина и участников. Каждая сессия состояла из 35 испытаний. Первые и последние 10 испытаний каждой тестовой сессии были испытаниями «разминка» и «охлаждение», в которых участнику были представлены только базовые звуки.Средние 15 испытаний были испытательными испытаниями, в которых все пять уровней акустической трансформации были представлены по одному разу для каждого из трех типов звука. Фаза тестирования состояла из 630 полных испытаний (180 испытаний на разминку, 270 испытаний и 180 испытаний на охлаждение). Каждый набор из трех сеансов обычно занимал 5–9 минут ( M = 6,32 мин), а вся фаза тестирования занимала 33–50 минут ( M = 40,23 мин).
После завершения этапа тестирования было проведено короткое интервью.Участникам был предоставлен список звуковых терминов, которые можно было использовать для описания имитируемых звуков свистка. Были предоставлены следующие рабочие определения: громкость (насколько громкий или мягкий звук в целом, в зависимости от амплитуды или интенсивности звука), высота звука (насколько высокий или низкий звук в целом, в зависимости от частоты звука), длина (как долго каждый звук — это общий звук, также называемый продолжительностью), тембр (свойство музыкальных тонов, которое позволяет отличать один инструмент от другого, когда высота и громкость остаются постоянными) и частотный контур (профиль частоты звука во времени).Затем экспериментатор продемонстрировал каждый звуковой словарный термин на слуховых примерах. Громкость была продемонстрирована последовательностью из трех чистых звуков тона, в которых звук увеличивался в объеме (высота и тембр оставались постоянными). Высота звука была продемонстрирована двумя чистыми звуками, воспроизводимыми фортепиано с постоянной громкостью (880 Гц против 220 Гц). Продолжительность была продемонстрирована с тремя нотами разной длины (1 с, 2 с, 4 с), сыгранными на рояле (высота и громкость оставались постоянными; сгенерированы в GarageBand версии 5.1, Apple Inc., 2009). Тембр был продемонстрирован с чистым тоном (средняя до) трех разных инструментов (валторна, труба и саксофон сопрано; громкость оставалась постоянной). Играли двумя разными свистками дельфинов, чтобы продемонстрировать разные частотные контуры (Discovery of Sound in the Sea, 2013). На листе звукового словаря отображались сопутствующие спектрограммы, показывающие изменения частоты во времени двух дельфиньих свистков. Наконец, эксперимент снова воспроизводил базовые звуки веревки, бутылки и мяча для участника и давал им имена перед воспроизведением каждого звука.Участникам разрешалось слышать любой из трех звуков столько раз, сколько они хотели.
В ходе интервью участников попросили описать различия, которые они слышали между базовыми звуками, используя словарные термины, описать различные сигналы, которые они использовали для идентификации каждого из трех объектов, и попытаться определить способы, которыми они слышали базовые звуки. были преобразованы во время фазы тестирования. Фаза интервью длилась 2–6 мин ( M = 4,03 мин). Весь эксперимент занял 54–76 минут ( M = 66.4 мин) до завершения.
Результаты и обсуждение
Показатели дельфина
Правильная пропорция (ПК) для испытаний на разминку и охлаждение (также известные как исходные испытания) была объединена. Базовый ПК был рассчитан для каждого типа звука и каждого измерения звука (всего три показателя ПК для каждого типа звука). Среднее значение PC для каждого типа звука в исходных условиях составляло 0,92, 0,96 и 0,96 для веревки, бутылки и мяча соответственно. Односторонний дисперсионный анализ не привел к значительным различиям между этими средними значениями, F (2,6) = 0.951, р. = 0,438. Для тестовых испытаний ПК был рассчитан для каждого из уровней измерений и отображается в формате. В среднем, небольшое снижение производительности произошло на уровнях представления как 120 дБ, так и 140 дБ. Однако это снижение не было статистически значимым, F (6,8) = 3,35, p = 0,059. SAY показал хорошие результаты независимо от разницы в громкости. Учитывая, что SPL свистков естественно меняется в зависимости от источника звука и из-за различий в дальности действия между сигнализатором и приемником, система распознавания, инвариантная по амплитуде, неудивительна.
Представление дельфинов.Пропорция правильная как функция (A) SPL (амплитуда), (B) центральной частоты и (C) продолжительности звука. Каждая серая точка данных представляет собой пропорцию, правильную для десяти испытаний, а черные точки данных представляют собой средние значения.
Полиномиальная модель, которая включала частотную транспозицию, тип звука и взаимодействие между ними, оказалась значимым предиктором ПК, F (8,10) = 26,76, p <0,001. Умеренное снижение PC произошло для CF = 11 кГц, а значительное снижение способности распознавания происходит для обоих сдвигов на ½ октавы.Вдобавок возникает интересная картина, в которой дельфин почти всегда выбирал мяч, когда центральная частота была на ½ октавы выше, чем базовые звуки (). Звуки веревки и бутылки имеют начальную частоту 8 кГц, а мяч - 10 кГц. Если дельфин взвешивал свое решение по стартовой частоте (то есть, если стартовая частота высокая, выбирайте мяч), то ошибочное решение по умолчанию использовать мяч в условиях высокой частоты можно объяснить. Конечно, правило «высокочастотного старта» не может объяснить все ошибки, допущенные дельфином, и требуются дополнительные механизмы принятия решений.Однако транспозиционная инвариантность, по-видимому, не является особенностью системы распознавания свистков этого дельфина.
Хотя продолжительность не оказала существенного влияния на ПК ( F (6,8) = 1,45, p = 0,303), стоит отметить характер ошибок. показывает, сколько раз дельфин выбирал не тот объект во время тестовых испытаний. Интересно, что когда продолжительность звука веревки была уменьшена до 250 мс, SAY почти каждый раз ошибочно выбирал бутылку. Поскольку звук веревки 250 мс очень похож на первую петлю в звуке базовой линии бутылки, дельфин, вероятно, классифицировал веревку 250 мс как одиночную петлю звука бутылки.
Таблица 3
Ошибки, допущенные участником-дельфином при каждом типе трансформации.Воспроизводимый звук отображается в левом столбце каждой матрицы, а выбор дельфина отображается в верхней строке. В столбце суммы указано общее количество неверных попыток для каждого типа звука
| AMPLITUDE | ||||||||||||
| веревка | бутылка | мяч | сумма | |||||||||
| Звук | веревка | 1 | 0 | 1 | ||||||||
| 1 | ||||||||||||
| 1 | ||||||||||||
| мяч | 0 | 5 | 5 | |||||||||
| 01071 | 0 0 0 0 0 0 0 0 | выбор | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| веревка | бутылка | мяч | сумма | 071 | 9111 9117 9117 9113 9117 9117 9117 12 | 16 | ||||||
| бутылка | 1 | 14 | 15 | |||||||||
| мяч | 4 | 7 | 0 0 0 9111 | |||||||||
| ПРОДОЛЖИТЕЛЬНОСТЬ | ||||||||||||
| 9111 9111 9111 9111 9111 9111 | 9111 | 9111 | 9117 9 1010 | мяч | сумма | |||||||
| звук | веревка | 14 | 1 | 15 | ||||||||
| бутылка | 3 | 3 | мяч | 0 | 4 | 4 |
отображает непосредственность распознавания с новыми тестовыми звуками, исследуя результат испытания, когда новый тестовый звук был впервые представлен дельфину.Квадратики с «Х» указывают на правильный выбор, а пустые поля указывают на неправильный выбор. Производительность первого испытания (также известная как переходный тест) важна, потому что не было возможности узнать правильный ответ из-за отзывов о предыдущих испытаниях. Все испытания после первого испытания могут быть изучены из-за положительного или отрицательного результата. Первое испытание было выше вероятности как для SPL, так и для продолжительности, но не превысило случайных характеристик для частотной транспозиции. Процент правильных результатов первого испытания составил 91.6% для SPL ( p <0,001, кумулятивный биномиальный тест, шанс = 0,33), 50,0% для частоты ( p = 0,063, кумулятивный биномиальный тест, вероятность = 0,33) и 83,0% для продолжительности ( p < 0,001, совокупный биномиальный тест, шанс = 0,33).
Таблица 4
Моментальность распознавания дельфином новых тестовых звуков.Знак «X» указывает на правильный выбор в самый первый раз, когда дельфину предъявили стимул, а пустое место указывает на неправильный выбор в первый раз.
| УЗД (дБ) | 120 | 125 | 135 | 140 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Трос | X | X | 0003 X | 0003 Бутылка | X | X | X | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Шарик | X | X | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| част.(кГц) | 7.07 | 9.00 | 11.00 | 14.14 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Трос | X | X | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Болл | X | X | X | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 010 00 | 250 | 400 | 600 | 1000 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Трос | X | X | 00 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| X | X | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ball | X | X | X | производительность0 9000 Сводное описание | 0 9000 кажется устойчивым к преобразованию по амплитуде и длительности; однако распознавание не распространялось на более частотные транспонированные звуки.Возможности человекаУчастники-люди очень точно распознавали типы звуков (). Точность характеристик для испытаний на разогрев и охлаждение (исходный уровень) во время фазы испытаний были объединены вместе, и было вычислено среднее значение для каждого типа звука. Средняя точность исполнения в исходных условиях для каждого типа звука составила 97,8%, 99,8% и 97,1% для веревки, бутылки и мяча соответственно. А 3 (группа: порядок A, B, C) x 3 (тип звука: веревка, бутылка, мяч) дисперсионный анализ (ANOVA), проведенный в испытаниях на разминку и охлаждение, выявил основной эффект группы; F (2, 27) = 3.36, p <0,05; и основной эффект звукового типа; F (2, 54) = 7,89, p <0,001. Характеристики звукового типа «бутылка» были значительно лучше, чем для звукового типа «веревка» и «мяч» (Newman-Keuls, , p <0,05). Участники порядка A (тот же порядок, что и дельфина) показали значительно лучшие результаты, чем участники порядка B ( M = 99,8% против 97,0%), но не было значительной разницы между порядками C и B или порядками A и C ( С = 97.8%, Newman-Keuls, p <0,05). Неясно, почему возник этот групповой эффект, но одна из возможностей состоит в том, что на выступления участников в исходных испытаниях мог повлиять их музыкальный опыт. Участники, случайно отнесенные к группе A, имели в среднем 7,5 лет музыкального опыта, тогда как участники, случайно отнесенные к группам B и C, имели в среднем 2,5 и 4,6 года опыта соответственно. Возможности человека.Пропорция правильная в зависимости от (A) SPL, (B) центральной частоты и (C) продолжительности звука.Каждая серая точка данных представляет собой пропорцию, правильную для десяти испытаний, а черные точки данных представляют собой средние значения. Обратите внимание, что шкала по оси Y отличается для и Рис. 6. Из-за почти безошибочного представления типа звука бутылки при испытаниях трансформации (и отсутствия дисперсии) он был исключен из всех ANOVA в сеансах испытаний трансформации. A 3 (группа: порядок A, B, C) x 2 (тип звука: веревка, мяч) x 5 (уровень преобразования: 60 дБ, 65 дБ, 60 дБ, 75 дБ, 80 дБ) ANOVA был выполнен на тесте амплитуды сеансы с последними двумя факторами в качестве повторных измерений.Не было основных эффектов группы, F (2, 27) = 2,81, p = 0,08, типа звука, F (1, 27) = 0,72, p = 0,40, или уровня трансформации, F (4,108 ) = 2,33, р = 0,06. Как и дельфин, люди-участники показали хорошие результаты независимо от амплитудных преобразований. A 3 (группа: порядок A, B, C) x 2 (тип звука: веревка, мяч) x 5 (уровень преобразования: 7,07 кГц, 9,00 кГц, 10,00 кГц, 11,00 кГц, 14,14 кГц) ANOVA был выполнен на частота сеансов тестирования с последними двумя факторами в качестве повторных измерений.Не было основных эффектов группы, F (2, 27) = 2,7583, p = 0,08. Был главный эффект типа звука, F (1, 27) = 10,56, p <0,01, и эффект взаимодействия между типом звука и группой, F (2, 27) = 3,47, p <0,05. Последующий анализ показал, что участники порядка C показали значительно лучшие результаты с типом звука веревки ( M = 92%), чем с типом звука мяча (M = 86%; Newman-Keuls, p <0.05). Участники, которые начинали и заканчивали тест с частотных сессий, демонстрировали отличную производительность от двух других групп. Не было основных эффектов трансформации, F (4, 108) = 2,12, p = 0,08. В отличие от дельфина, у участников-людей не было снижения производительности ни на одном уровне частотного преобразования. A 3 (группа: порядок A, B, C) x 2 (тип звука: веревка, шар) x 5 (уровень преобразования: 250 мс, 400 мс, 500 мс, 600 мс, 1000 мс) ANOVA был завершен на продолжительность тестовых сессий с последними двумя факторами в качестве повторных измерений.Был обнаружен основной эффект группы, F (2, 27) = 4,31, p = 0,02. Последующий анализ показал, что группа A ( M = 99,8%) показала значительно лучшие результаты, чем группа B ( M = 91,3%). Не было значительных различий между группой A и группой C ( M = 94,3%) или группой B и группой C (Newman-Keuls, p <0,05). Участники, получившие сеансы продолжительности после двух других типов трансформации (в середине фазы тестирования), показали значительно лучшие результаты, чем те, кто получил сеансы продолжительности в начале и в конце фазы тестирования.Был найден основной эффект звукового типа: F (1, 27) = 4,64, p = 0,04. Последующий анализ показал, что эффективность веревки со звуком ( M = 96,1%) была значительно лучше, чем с точки зрения со звуком мяча ( M = 94,2%; Newman-Keuls, p <0,05). Наконец, был обнаружен основной эффект трансформации: F (4, 108) = 4,19, p <0,01. Последующий анализ показал, что производительность при преобразовании 250 мс ( M = 91.4%) был значительно хуже, чем другие уровни преобразования (400 мс: M = 96,1%, 500 мс: M = 96,4%, 600 мс: M = 96,9%, 1000 мс: M = 95,0 %; Neumann-Keuls, p <0,05). Падение производительности на 250 мс отражает производительность дельфина. Участники-люди совершили ошибки, отличные от ошибок дельфинов (,). Чаще всего люди путали веревку и мяч, а дельфин — бутылку и мяч или веревку и бутылку.Люди-участники обнаружили, что звуковой тип бутылки очень отличен (три коротких звука) по сравнению с типами звука веревки и шара (каждый звук по отдельности), и поэтому почти никогда не ошибались в звуках бутылки. Сравнение ошибок (путаницы между типами звука), сделанных участниками-людьми и дельфином для каждого преобразования звука. Таблица 5Ошибки, допущенные участниками-людьми ( N = 30) для каждого типа преобразования.Воспроизводимый звук отображается в левом столбце каждой матрицы, а выбор участника отображается в верхней строке.В столбце суммы указано общее количество неверных попыток для каждого типа звука.
Ошибки, допущенные участниками при преобразовании амплитуды, в основном касались веревки и мяча (41 ошибка, 87% ошибок).Небольшая часть ошибок была сделана между бутылкой и мячом (4 ошибки, 9% ошибок) и веревкой и бутылкой (2 ошибки, 4% ошибок). Тип звука шара обычно ошибочно принимался за тип звука веревки, на него приходилось 23 из 41 ошибки, в основном возникающих при преобразовании 75 дБ. Кроме того, тип звука веревки был перепутан со звуком мяча почти одинаково на всех уровнях трансформации, всего 18 ошибок. Наибольшее количество ошибок участники-люди были допущены во время частотных сессий.Всего было допущено 133 ошибки, в основном между веревкой и мячом (123 ошибки, 92%). Небольшая доля ошибок была сделана между бутылкой и мячом (8 ошибок, 6%) и веревкой и бутылкой (2 ошибки, 2%). Тип звука мяча путали с типом звука веревки чаще всего на всех уровнях трансформации (включая базовый уровень), что составляет 76 из 123 ошибок веревки и шара. Звуковой тип веревки реже путали с типом звука шара, но на него приходилось 48 из 123 ошибок веревки и шара. Ошибки были распределены почти равномерно по всем уровням трансформации, за исключением базового уровня 1000 Гц, где тип звука веревки был перепутан с типом звука мяча только дважды. В общей сложности 91 ошибка была сделана во время тренировок, в основном между скакалкой и мячом (79 ошибок, 87%). Такая же пропорция ошибок была сделана между бутылкой и мячом и веревкой и бутылкой (6 ошибок или 7% каждая). Тип звука мяча больше всего путали с типом звука веревки, на него приходилось 49 из 91 ошибки, причем наибольшее количество ошибок (17) приходилось на преобразование 250 мс. Тип звука веревки был принят за второй по частоте тип звука мяча, на него приходилось 30 из 91 ошибки.Большинство этих ошибок произошло при экстремальных преобразованиях, наиболее удаленных от базового звука, 250 мс (7 ошибок) и 1000 мс (9 ошибок). Первое испытание с участием людей показано на. Процент участников, которые ответили в первом испытании правильно на любой тип звука и трансформацию, варьировался от 77% до 100%. В условиях, при которых наименьшее количество участников-людей правильно ответило на первое испытание [мяч при 900 Гц (77%), веревка при 900 Гц (80%), мяч при 400 мс (83%)], дельфин был правильным в первом испытании. испытание в аналогичных условиях.В условиях, когда дельфин был неправильным в первом испытании, высокий процент участников-людей ответили правильно. Таблица 6Непосредственность распознавания с новыми тестовыми звуками для участников-людей и дельфинов.Жирный текст указывает на то, что дельфин был правильным при первом испытании, в то время как обычный текст указывает на то, что дельфин был неправильным при первом испытании. Показан процент участников-людей (N = 30), которые получили правильный ответ в первом испытании для каждого типа звука и преобразования.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||