| нет фото | |||||
| ВозникновениеOrigin | Создатель, первоносительFounder, first-bearers | ЗаконодателиLawgivers | ПотребителиUsers | ПроизводныеDerivatives | |
| Грамматический статусGrammatical status | РейтингRating | Ресурсность, распределениеResourceness, allocation | Релевантность, престижностьRelevance, prestige | СтатистикаStatistics | |
| Лингвистический статусLinguistic status | утор — м. | Архивы: фото, видео, аудиоArchives: photo, video, audio | |||
| Специфический смыслSpecific meaning | ХронологияChronology | ||||
| Подразделения, морфемыSubdivisions, morphemes | Этимология, историяEtymology, history | ||||
| Человеческая средаHuman environment | География распространенияGeography of spreading | ||||
| Разночтения, разновидности, разносмыслыSpellings, varieties, dissents | Социология распространенияSociology of spreading | ||||
| АналогиAnalogues | ФетишизацииFetishizations | ||||
| Медиа, новостиMedia, news | Переводы вовнеTranslations outside | ||||
| Лица причастныеPersons participial | Замены внутриSubstitutions inside | ||||
| Нормирования словарныеRegulation lexicographical | ОтзывыOpinions | ||||
| Фонемоциды, мутацииPhonemacides, mutations | Пословицы, крылатые словаProverbs, pithy sayings | ||||
| Оборотничество, лжесмыслReverseness, false meanings | БиблиографияBibliography | ||||
| ЗабвениеOblivion | СетьWeb | ||||
 нарезка в конце деревянного сосуда — бочки, кадки и т.п. — для вставки дна. утор — м. нарезка в конце деревянного сосуда — бочки, кадки и т.п. — для вставки дна.
нарезка в конце деревянного сосуда — бочки, кадки и т.п. — для вставки дна. утор — м. нарезка в конце деревянного сосуда — бочки, кадки и т.п. — для вставки дна.Выдержка вина в бочке: плюсы и минусы, прошлое и будущее
Когда вы всерьез увлекаетесь темой вина, вас начинает раздражать вкус новой дубовой бочки в бокале. Сомелье презрительно фыркают, уловив эти нотки в вине, и заявляют, что такое может понравиться только тем, кто ничего не смыслит в хорошем напитке.
Сомелье презрительно фыркают, уловив эти нотки в вине, и заявляют, что такое может понравиться только тем, кто ничего не смыслит в хорошем напитке.
В 90-е и даже еще в начале 2000-х ничего подобного не говорили. Тогда наличие в вине аромата новой дубовой бочки в сочетании с нотками переспелых фруктов только поощрялось. Сегодня мода на вино изменилась, и сомелье не любят «дубовые» вина, а самые продвинутые виноделы отказываются от использования новых бочек на производстве. Вино зреет у них в старых бочках, в популярных сегодня квеври (конусообразные керамические амфоры, традиционные для Кавказа) или даже в огромных керамических сферообразных емкостях.
Отказ от использования новых дубовых бочек для выдержки вина кажется вполне разумным. Какой смысл в тщательном поиске удачного места для виноградника и последующем внимательном выращивании винограда, если в итоге в вине не будет того самого уникального терруара, а останется лишь перебивающий всё вкус и аромат бочки?!
Но не стоит быть столь категоричными: дубовые бочки нужно использовать с умом, далеко не все они так плохи, как кажется. Вообще идея использовать дуб для придания вину тех или иных характеристик довольно нова. Изначально бочки использовали в виноделии просто потому, что это было доступно и удобно, они были прочнее терракоты и легче в транспортировке.
Вообще идея использовать дуб для придания вину тех или иных характеристик довольно нова. Изначально бочки использовали в виноделии просто потому, что это было доступно и удобно, они были прочнее терракоты и легче в транспортировке.
В 1990-х годах мир любителей вина пристрастился ко вкусу, который приобретает вино, выдерживаемое именно в новых дубовых бочках. В таком вине отчетливо ощущается вкус и аромат кокоса и ванили, ореховые нотки, копченые или жареные тона.
К преимуществам выдержки в бочке относят контакт вина с кислородом, который происходит благодаря структуре дерева, и которого лишена, например, выдержка в чанах из нержавеющей стали. Также плюсом считается наличие на дне бочки осадка из мертвых дрожжевых частиц, которые придают вину особые вкусо-ароматические характеристики. Такое вино называют «выдержанным на осадке» («sur lie» по-французски).
Бургундия и бочка
Однако даже самые продвинутые и модные сомелье не могут устоять перед великолепными красными и белыми винами из Бургундии: непревзойденными образцами элегантности и силы. Здесь в любом погребе, особенно самых топовых виноделен, вы обязательно найдете небольшие дубовые бочки.
Лучшие виноделы Бургундии являются приверженцами такого понятия, как élevage (фр. воспитание). Это целый набор определенных навыков, которые позволяют делать действительно превосходные вина. И одним из важнейших средств такого воспитания вина является бочка.
Традиционно красные бургундские оставляют в небольших дубовых бочках на зиму, а весной, когда погреб прогревается, у вина в бочках начинается второе яблочно-молочное брожение. По окончании брожения вино остается в бочках, пока не стабилизируется и не станет прозрачным.
Лучшие белые вина Бургундии ферментируются в бочках, после чего винодел принимает важные решения: перемешивать ли вино в бочке, переливать ли его в другую бочку и сколько еще времени вино должно провести в дубе…
Всякий раз, когда виноделы в других странах пытаются сделать высококачественные версии бургундского пино нуар и шардоне, они используют небольшие дубовые бочки. Когда вы ферментируете вино в новой бочке, химический процесс может придать вину определенные ароматы. В результате, как ни странно, получаются гораздо менее «дубовые» вина, чем если бы вы ферментировали шардоне сначала в стальном резервуаре, а затем переливали его в бочку.
Когда вы ферментируете вино в новой бочке, химический процесс может придать вину определенные ароматы. В результате, как ни странно, получаются гораздо менее «дубовые» вина, чем если бы вы ферментировали шардоне сначала в стальном резервуаре, а затем переливали его в бочку.
Современная реакция на вина, выдержанные в бочке, не ошибочна, но она неоднозначна. Модные сомелье разлюбили бочки, потому что многие производители злоупотребляли дубовой выдержкой в погоне за пародийными винами «аля Бургундия». Но бочки действительно имеют место в создании некоторых классических винных стилей, которые знают и любят даже самые взыскательные профессионалы и энтузиасты виноделия.
Источник: Vinepair
Перевод: Марина Кирсанова
Вам может понравиться
адјутор
УТОРЁННЫЙ — УТОРЁННЫЙ, уторённая, уторённое; уторен, уторена, уторено (обл.
УТОРЁННЫЙ — УТОРЁННЫЙ, уторённая, уторённое; уторен, уторена, уторено (обл.). прич. страд. прош. вр. от уторить. Толковый словарь Ушакова. Д.Н. Ушаков. 1935 1940 … Толковый словарь Ушакова
уторённый — уторённый; кратк. форма ён, ен а (от утор ить) … Русский орфографический словарь
УТОР — УТОР, утора, чаще мн., муж. (спец.). Нарезка, вырезка в конце деревянного сосуда для вставки дна. Нарезать уторы в бочке. Толковый словарь Ушакова. Д.Н. Ушаков. 1935 1940 … Толковый словарь Ушакова
утор — сущ., кол во синонимов: 1 • нарезка (10) Словарь синонимов ASIS. В.Н. Тришин. 2013 … Словарь синонимов
уторік — прислівник незмінювана словникова одиниця … Орфографічний словник української мови
утор днища бака — кольцевой паз днища бака — [А. С.Гольдберг. Англо русский энергетический словарь. 2006 г.] Тематики энергетика в целом Синонимы кольцевой паз днища бака EN tank bottom chime … Справочник технического переводчика
С.Гольдберг. Англо русский энергетический словарь. 2006 г.] Тематики энергетика в целом Синонимы кольцевой паз днища бака EN tank bottom chime … Справочник технического переводчика
утор днища резервуара — — [http://slovarionline.ru/anglo russkiy slovar neftegazovoy promyishlennosti/] Тематики нефтегазовая промышленность EN tank bottom chimetank bottom chime … Справочник технического переводчика
утор — нарезка в клепках бочки, которая держит дно , укр. утори (мн.) – то же, болг. вътор край, утор , сербохорв. у̏тор, словен. votòr, род. п. tȯra, чеш. utor, слвц. utor, польск. wątor, в. луж. wutora, н. луж. hutora. Праслав. *ǫtorъ, с… … Этимологический словарь русского языка Макса Фасмера
уторённый — ён, ена уторённый, ён, ена (от уторить) … Словарь употребления буквы Ё
Утор — м. Нарезка в конце деревянного сосуда бочки, кадки и т.п. для вставки дна. Толковый словарь Ефремовой. Т. Ф. Ефремова. 2000 … Современный толковый словарь русского языка Ефремовой
Толковый словарь Ефремовой. Т. Ф. Ефремова. 2000 … Современный толковый словарь русского языка Ефремовой
Розлив пива в бочки
Транспортные бочки для пива — это деревянная или металлическая тара емкостью 25, 50, 100 и 200 л, наиболее распространены бочки ёмкостью от 25 до 100 л, в то время как бочки большего объема используются только для поставок пива крупным центрам сбыта. В некоторых случаях встречаются бочки емкостью 12,5 л (0,12 гл).
Транспортные бочки имеют стандартную форму. Отверстие для наполнения, служащее одновременно и отверстием для мойки, находится в дне бочки. Оно снабжено чугунным или бронзовым кольцом и закрывается деревянной пробкой (шпунтом) диаметром 44-47 мм. Часто в Чехословакии, главным образом у металлических бочек, кольцо имеет нарезку и закрывается ввинчивающейся металлической пробкой, герметичность которой обеспечивается за счет дополнительной прокладки. Второе отверстие бочки, предназначенное для разливного крана, размещается в одном из доньев, ближе к периметру и закрывается корковой или деревянной пробкой (40×40 мм).
Нижняя узкая часть этого затвора, предназначенная для введения иглы разливного крана вплоть до противоположного дна бочки, тоже закрывается небольшой корковой пробкой диаметром 16-18 мм. Снаружи затвор защищен от загрязнений алюминиевой заслонкой. На каждой транспортной бочке должны быть данные о действительном ее объеме с датой маркировки, номером бочки и названием предприятия (завода), который выпустил пиво.
Деревянные бочки
В настоящее время в Чехословакии пользуются транспортными бочками для пива, изготовленными из колотой или резаной дубовой клепки. Преимуществом деревянных бочек является то, что они хорошо защищают пиво от резких колебаний температуры, которая выравнивается в них до температуры окружающей среды только спустя 24 ч. Наблюдающийся в последнее время отказ от деревянных бочек не связан только с недостатком качественной дубовой клепки; главной причиной являются недостатки этих бочек по сравнению с алюминиевыми. Прежде всего они слишком тяжелые (их масса в 3 раза больше, чем алюминиевых оочек) и это повышает транспортные расходы и затрудняет операции при погрузочно-разгрузочных работах. Уход за деревянными бочками сложен и дорог. Кроме обычной мойки перед использованием в них надо менять внутреннее смоляное покрытие Это требует специального оборудования для рассмоления, смоления и обкатки просмоленных бочек.
Прежде всего они слишком тяжелые (их масса в 3 раза больше, чем алюминиевых оочек) и это повышает транспортные расходы и затрудняет операции при погрузочно-разгрузочных работах. Уход за деревянными бочками сложен и дорог. Кроме обычной мойки перед использованием в них надо менять внутреннее смоляное покрытие Это требует специального оборудования для рассмоления, смоления и обкатки просмоленных бочек.
Поврежденные бочки должны ремонтировать бондари — специалисты высокой квалификации. Не только при ремонте и смолении, но и при обычном использовании деревянная бочка несколько меняет форму, а следовательно, и объем, в результате бочки следует заново перемерить и перемаркировать согласно требованиям стандарта. Кроме того, внутренность бочек трудно содержать в необходимом с биологической точки зрения состоянии. Даже новое смоление не гарантирует практической стерильности, поскольку смоляное покрытие при розливе пива легко растрескивается от противодавления.
Стремясь сэкономить дубовую клепку и снизить массу бочек в Чехословакии испытывали бочки из буковой клепки За границей испытывали бочки из прессованных опилок, пропитанных полиамидном смолой.
Металлические бочки
Металлические транспортные бочки в качестве тары начали использовать для экспортного пива в начале столетия Эти первые бочки были стальные, эмалированные или лакированньге внутри. Их форма была цилиндрической и сдвоенные стены образовывали изоляционную воздушную рубашку. Для транспортировки по морю их помещали в деревянную упаковку, чтобы ограничить резкие изменения температуры пива. За рубежом металлические бочки стали применять после второй мировой войны, в некоторых странах они полностью заменили деревянные бочки. В Чехословакии металлические бочки стали широко применять только в последние годы.
Металлические бочки с простыми стенками, похожие на деревянные, начали изготавливать из стали и смолить внутри. Их недостаток заключался в том, что смоляная поверхность от ударов легко отлетала. В настоящее время преобладают транспортные бочки из алюминиевых сплавов (внутренний слой из чистого алюминия). Они прочнее применявшихся ранее бочек из чистого алюминия и устойчивее к ударам. Внутренняя поверхность из чистого алюминия анодирована, чтобы лучше противостояла коррозии. Алюминиевые транспортные бочки сваривают из двух половин. Их качество в значительной степени зависит от выполнения сварки. Нарушение герметичности чаще всего наблюдается в швах у выпускного отверстия, которое больше всего страдает при транспортировке.
Внутренняя поверхность из чистого алюминия анодирована, чтобы лучше противостояла коррозии. Алюминиевые транспортные бочки сваривают из двух половин. Их качество в значительной степени зависит от выполнения сварки. Нарушение герметичности чаще всего наблюдается в швах у выпускного отверстия, которое больше всего страдает при транспортировке.
Невозможно абсолютно исключить коррозию алюминиевых бочек. Она появляется от мойки щелочными средствами и главным образом от прокисших остатков пива. Поэтому внутри бочки покрывают стойкими синтетическими покрытиями (алупласт, дуропласт). В последнее время за рубежом появились транспортные бочки из пластических масс, главным образом из полиэтилена (дуролен, бакелор и т. д.). Они легкие, очень прочные и практически их можно мыть всеми моющими средствами. Большого опыта в работе с ними пока нет.
Деревянные бочки изготовление в домашних условиях. Как сделать деревянную бочку своими руками чертежи
С древних времен вино и коньяки вызревали в деревянных бочках, технология производства которых была засекречена.
Сегодня бочки большого размера укомплектовываются дубовыми пробками, имеют удобную подставку и оснащены краном.
Археологические раскопки позволяют говорить о существовании деревянных бочек задолго до начала нашей эры. Уже тогда люди убедились в непревзойденных качествах именно бочек из дерева. Они были прочнее утвари, изготовленной из керамики и глины, и служили дольше. К тому же менялся вкус алкогольных напитков, выдержанных в деревянных бочках, в лучшую сторону.
Придерживаясь технологии при изготовлении бочек, можно существенно продлить им жизнь.
Наши специалисты придерживаются всех разработанных технологий и используют секреты при производстве деревянных бочек из дуба.
ПРОИЗВОДСТВО ДУБОВЫХ БОЧЕК
Прежде, чем начать производство деревянных бочек, нужно выбрать качественный материал: ствол старого дуба, возрастом от 80 лет и старше. Заготовка дерева производится только в зимний период. Стволы не сразу забирают на производство. Сначала дают дереву высохнуть (сушка проходит от 2,5 до 3 лет), после распиливают на нужные отрезки вдоль волокна.
Далее, на высококачественном оборудовании создаются клепки, которые являются основанием любой бочки. Форма у них оригинальная: у торцов клепка уже и толще, нежели в средине, что позволяет придать гнутую форму бочке. Получившиеся заготовки отправляют в сушильную печь на три недели.
Для того чтобы избежать протечек изделия и минимизировать испарение спирта во время выдержки используется радиальный распил клепки.
Готовые и хорошо просушенные клепки укладывают в пирамидки.
На следующем этапе изготовления бочек собирают остов из рабочих обручей. Обруч прикрепляют к клёпкам, расставленным на правильном расстоянии друг от друга. Обычно используют три клёпки. Затем в промежутки между ними устанавливают следующие клёпки, прижимая их максимально плотно. Главный обруч опускают вниз, одновременно добавляя срединный обруч, а далее, сняв зажимы, устанавливают торцевой обруч.
В процессе производства используют металлический трос, с помощью которого стягивают клёпки, чтобы пресечь появление даже минимальных щелей.
Чтобы добиться плотности бочки, не пропускающей воздуха совсем, внутри неё разводят костёр. Дно на этом этапе не прикреплено.
Обжиг бочки происходит не менее получаса.
Для некоторых бочек применяют обжиг с помощью опилок. Подкаливая древесину, чтобы она приобрела красивый золотистый цвет.
На последнем этапе вставляются днища, изготовленные из тех же клёпок, плотно подогнанных и прифугованных друг к другу. Далее бочка шлифуется и на ней устанавливаются постоянные обручи, изготовленные из оцинкованной или нержавеющей стали толщиной в 2 мм.
Готовые изделия упаковываются в термоусадочную пленку, чтобы обеспечить сохранность груза при перевозке. В этом же виде товар доходит до конечного потребителя.
УСЛУГИ НАШЕГО МАГАЗИНА
Под торговым брендом «БОНПОС» производятся высококачественные бондарные изделия. На нашем сайте представлены дубовые бочки и кадки от 3-х до 200 литров. Визуально все изделия оформлены стильно.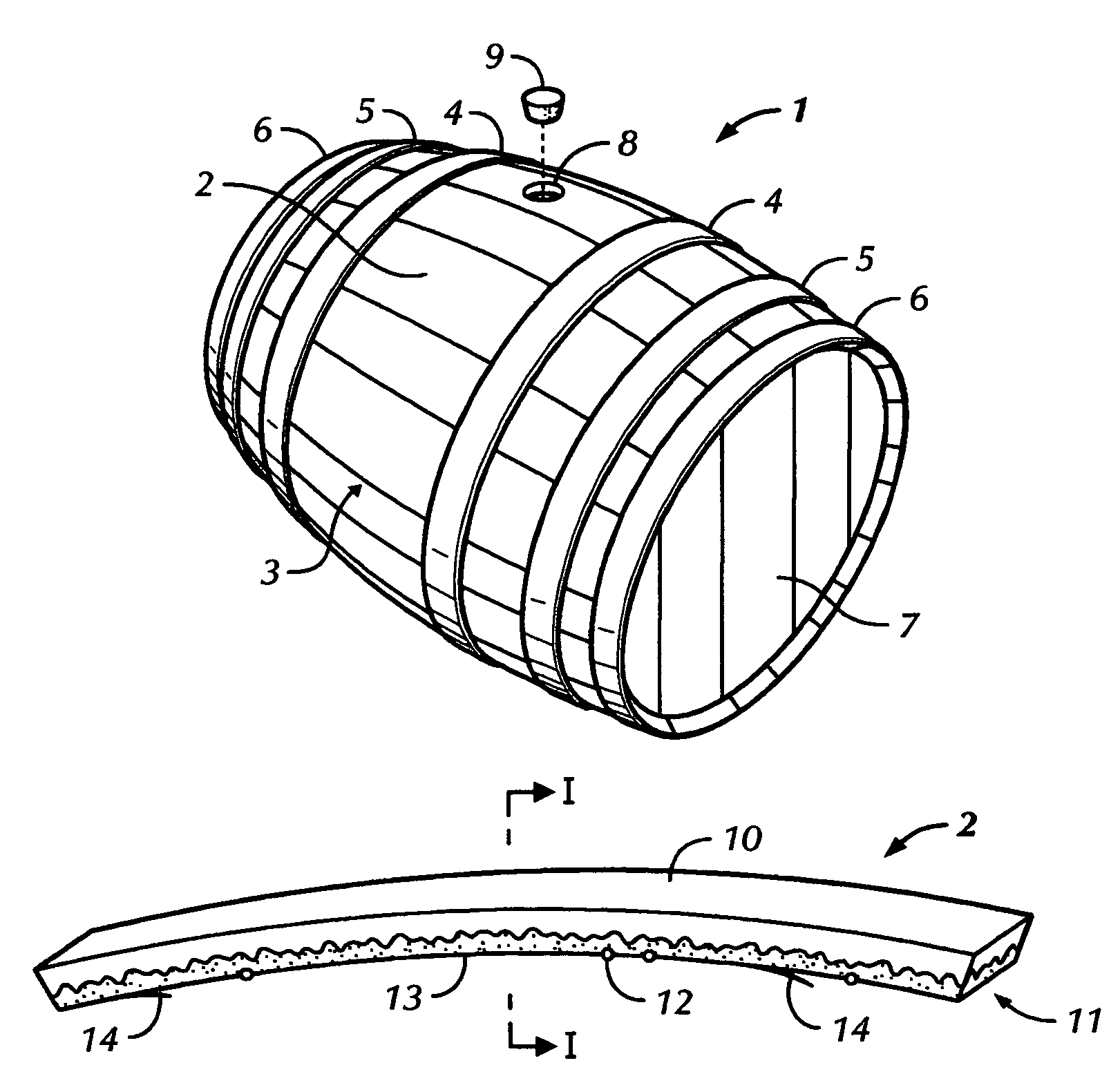 Качество наивысшее. Они не только выполняют свои прямые функции, но также и украсят ваш интерьер.
Качество наивысшее. Они не только выполняют свои прямые функции, но также и украсят ваш интерьер.
(Last Updated On: 19.09.2017)
Как сделать бочку из дерева своими руками в современном обществе? Наверняка, сегодня не придётся выдалбливать изделие из цельного ствола дерева, как это было раньше. Современная бочка сильно отличается от своих собратьев-предшественников. Важнейшим шагом к её изготовлению послужило появление металлических обручей, которые очень крепко стягивали конусную посуду.
Из какой древесины лучше изготовить бочку
Дубовое дерево
Превосходно колется и при распаривании приобретает необыкновенную гибкость. Именно это качество очень ценится. Но, самое главное, в дубовой древесине находятся консервирующие вещества под названием тиллы, которые предохраняют дерево от гниения. Поэтому продукты в дубовых бочках могут сохранять свои свойства на протяжении многих десятков лет.
Кедр
Также используется при изготовлении бочек. По своим свойствам он мягкий, гибкий и лёгкий. Раньше считали, что в самом кедре содержатся вещества, которые уничтожают микробов. Поэтому можно сделать бочку из кедра своими руками, в которой напитки будут храниться очень долгое время и не портиться.
Раньше считали, что в самом кедре содержатся вещества, которые уничтожают микробов. Поэтому можно сделать бочку из кедра своими руками, в которой напитки будут храниться очень долгое время и не портиться.
Можжевельник
Внутри ствола имеет красно-коричневую древесину, а заболонь намного светлее, желтоватого цвета. Древесина этого дерева характерна плотностью, крепостью и тяжёлым весом, причём, очень удобна в обработке – хорошо режется, не трескается.
Древесина сосны
Имеет средние показатели – невысокую твёрдость и среднюю прочность, высокую упругость, хорошо гнётся. Из-за специфического смоляного запаха в сосновых бочках не хранят пищевые продукты.
Как смастерить бочку в домашних условиях
Вопрос как сделать бочку из дерева своими руками волнует многих мастеровых людей, которые хотели бы овладеть новыми секретами работы с деревом.
Для изготовления бондарных изделий применяются клёпки или лады. Это прямоугольные деревянные дощечки, полученные распиливанием комля или ствола дерева на отдельные части. Другой способ – это раскалывание колод или чурбаков.
Другой способ – это раскалывание колод или чурбаков.
Пиленые лады получаются очень прочными. Колотые сделать сложнее. Самое главное – это уметь расколоть чурку так, чтобы клёпки получились чистые и ровные, и отходов щепы оставалось немного.
Чтобы сделать бочку из дерева своими руками необходимо правильно расколоть деревянный чурбак в направлении радиуса, желательно при этом попасть в сердцевину. Если топор ударил чуть левее или правее сердцевины, то линия скола называется тангентальной. В этом направлении почти все породы деревьев колются намного тяжелее.
Для изготовления клёпок пойдут и обыкновенные плахи. Из них нужно выбирать только те, где слои расположены вдоль, а не поперёк. Готовые лады просушивают и аккуратно складывают.
Как изготовляют обручи для стяжки
Железные обручи начали применять очень давно. Первоначально ими стягивали вёдра, состоящие из шестнадцати клёпок. Причём для крепости их дополнительно пробивали гвоздями.
В настоящее время обручи вырезают из прочной листовой стали, затем кузнецы украшают их всевозможными штампованными рисунками. Готовые обручи покрывают битумным лаком или олифой. Если слой олифы просушить паяльной лампой, получится красивый светло-коричневый цвет.
Готовые обручи покрывают битумным лаком или олифой. Если слой олифы просушить паяльной лампой, получится красивый светло-коричневый цвет.
Как собрать коническую основу бочки
Сначала постараемся соединить подготовленные боковые клёпки и временно стянуть их постоянными обручами. Для этого к одному из обручей (лучше взять малого диаметра) прикрепите две-три начальные клёпки. Аккуратно придерживая каждую вставляемую дощечку рукой, заполняем всё пространство недостающими клёпками. Особенно осторожно нужно вставить последнюю клёпку. Может получиться так, что для неё останется мало места. Возьмите и подстругайте её немного, ровно присоединив к остальным клёпкам. Такая работа, обычно, выполняется с двумя, тремя помощниками.
Схема сборки бочки:
С помощью молотка и деревянной выколотки нужно осадить обруч так, чтобы он плотно сел на основу бочки. Так же осаживается обруч большего диаметра.
Последняя операция – установка днища. Его аккуратно ставят в паз, переворачивают бочку и закрепляют большой обруч. Если соединение получилось крепким и надёжным, то временные обручи меняют на постоянные.
Если соединение получилось крепким и надёжным, то временные обручи меняют на постоянные.
Теперь вы представляете, как сделать бочку из дерева своими руками, используя только тот материал, который имеется у каждого в хозяйстве.
Люди, занимающиеся производством собственных вин, а то и просто солением огурцов, прекрасно знают — нет лучшей тары, чем деревянная бочка. Почему? Во-первых, дерево — экологически чистый материал, во-вторых, вина содержат спирт, значит, взаимодействуя с синтетическими материалами — пластиком или капроном, он может растворить химический состав тары, и она смешается с компонентами вина.
Процесс производства дубовой бочки не сложный, но он требует четкого соблюдения инструкции, аккуратности и внимательности.
Купить бочку не проблема, однако если есть желание и владелец домашней винной фабрики радеет за качество, то деревянная бочка своими руками будет главным доказательством, что вино и коньяк сделаны с применением правильной технологии. Кроме того, изготовление деревянных бочек — процесс трудоемкий и долгий, но без премудростей.
Этапы проведения работ
Дуб, ясень и вишня считаются наиболее подходящими деревьями для производства из них бочек.
Общеизвестно, что самым надежным материалом для изготовления деревянных бочек будет дуб. Он подойдет как для вин и коньяков, так и для солений. Дуб — природный антисептик, закись и плесень там не возникнет. Но если с этим проблема, можно поменять дуб на вишню или ясень — в этих деревьях содержится синильная кислота, которая тоже не даст развиться болезнетворным организмам, хотя их свойства слабее, чем у дуба. Не нужно бояться, что кислота попадет в продукты: во-первых, бочка тщательно вымачивается перед заливкой вин, а во-вторых, какой-то процент синильной кислоты содержится и в самих винах, организму он не опасен.
Расчет древесины. Доски, носящие название клепки, имеют двояковыпуклые стороны для придания бондарному изделию выпуклости. Чтобы их сделать такими, нужно взять нижнюю часть ствола дерева и расколоть подобием рубки дров. Если его аккуратно пилить, то нарушится природная целостность волокон, что плохо для такого изделия. Сразу приступать к фигурному выпиливанию не стоит — поленья нужно просушить в течение 2 месяцев. Причем сушить не под палящим солнцем, а в темном прохладном помещении.
Сразу приступать к фигурному выпиливанию не стоит — поленья нужно просушить в течение 2 месяцев. Причем сушить не под палящим солнцем, а в темном прохладном помещении.
Обработка поленьев для будущих бочек делается с помощью рубанка. Придавая им нужную форму, верх и низ нужно делать толще середины. Насколько — решит хозяин. Обычно это 1,5 см. Движения рубанком усиливаются к середине полена, тогда можно получить такую форму, как у классических бочек. В основном форму бочкам можно придать любую — пузатую, колодную, трапециевидную. Главное, все сделать правильно.
Подготовка скрепляющих обручей. Они могут быть как железными, так и деревянными. Деревянные имеют преимущества по красоте, защите от коррозии, однако не так прочны, как железные, а ведь хозяину винного погреба будет важно сохранить вина и бочки, в том числе от механического повреждения. Поэтому железо предпочтительнее. Итак, нержавеющая сталь режется на полоски, для лучшего скрепления ее можно проковать в местах соединения друг с другом. После этого на концах пробиваются отверстия для гвоздей, которые будут играть роль заклепок.
После этого на концах пробиваются отверстия для гвоздей, которые будут играть роль заклепок.
Набивка готовых обручей и подготовка остова. К малым в диаметре обручам с помощью фиксаторов цепляют три подготовленные доски. Расположить их нужно на одинаковом расстоянии друг от друга. Если расчет был верен, оставшиеся доски войдут впритык. После полной вставки, используя молоток и насадку, следует осадить обручи до того момента, когда их уж нельзя будет снять. Постукивая с противоположных концов, можно добиться хорошего эффекта.
Но нельзя переусердствовать: приготовленные долгим способом доски треснут, процесс придется начинать заново. Также треснуть подготовленные плашки могут и от долгого пересушивания. Так подготавливается один край бочки. Обруч большего диаметра нужно надеть сразу после первого края, действуя молотком. Теперь очередь второго края: заготовка перед монтажом второго конца долго парится. Делается это для того, чтобы дерево стало мягким и податливым. После этого заготовку переворачивают раскрытой стороной вверх и на распаренное дерево накидывают веревку, которую скручивают, чтобы концы клепок сошлись. Не отпуская веревки, нужно надеть обруч. Такая работа не делается в одиночку — кто-то скручивает веревку, а кто-то надевает обруч.
Завершение работы
Когда остов готов, его закаливают.
Способов много, в основном это обжиг. При этом запах и терпкость жженого дерева передастся вину, что тоже будет оценено. Делается это так: внутрь на бок готового остова кладут стружку какого-либо плодового дерева — предпочтительнее вишня. Поджигают и медленно перекатывают с боку на бок для равномерного тления. Костров внутри заготовки разводить не надо, по неопытности можно и спалить изделие. Нельзя применять никаких жидкостей для розжига — они в своем составе имеют химические элементы, которое дерево впитает. Можно применить паяльную лампу, но с осторожностью — сильный огонь может оставить в остове незамеченные тления, отчего изделие сгорит.
Завершающим этапом создания бочки считается ее шлифовка снаружи, высверливание отверстий для залива и прорезание желоба для доньев.
Далее процесс изготовления бочки заключается в следующем: заготовку обрабатывают инструментом — обрезают неровные концы, шлифуют снаружи, высверливают отверстие для залива и прорезают желоба для доньев. Днища бочек представляют собой два круга, вырезанные из подобия щитов. Щиты готовятся так: сколачиваются доски внахлест и скрепляются дополнительно скобами. Круги затачиваются так, чтобы их края уходили в скос. Нужно точно рассчитать диаметр доньев. Это делается после окончательной сборки остова, потому что сразу этого сделать не получится. Затем днища вставляются, посредством ослабления крайних обручей. После вставки одного нужно снова набить обруч, затем точно так же поступить и со вторым дном.
Проверка на герметичность. Естественно, бочка проверяется жидкостью, то есть водой. Она первое время даст течь, пока дерево не разбухнет. Отводится времени на то, чтобы бочка перестала протекать, не более часа. Если же она все равно течет, нужно найти щель и заделать ее. Бондари используют для таких целей стебли камыша. Их затыкают между клепками острым и тонким предметом, к примеру, ножом. Кроме того, покрыть бочку можно снаружи — воском. Только нужно взять действительно пчелиный — продукты не должны быть с химией.
Их затыкают между клепками острым и тонким предметом, к примеру, ножом. Кроме того, покрыть бочку можно снаружи — воском. Только нужно взять действительно пчелиный — продукты не должны быть с химией.
После того как изготовление бочки практически закончено — она закалена, обожжена и проверена на герметичность, ее парят или дают отстояться с водой, чтобы из дерева ушли все посторонние запахи, которые может перенять и вино. Можно обработать внутреннюю поверхность бочки перекисью водорода или марганцовкой.
Подведение итогов
Бочку своим руками сделать несложно, если внимательно следовать рекомендациям. Уход за ней крайне прост — нужно регулярно ее пропаривать или кипятить, а затем высушивать очень тщательно, если ее применение откладывается.
Еще одно важное правило: если бочка предназначена для жидких продуктов — рассолов, алкоголя или воды, то ее нельзя надолго оставлять сухой. Если же она применяется для складирования сухих продуктов, то ее нельзя мочить.
Не правда ли, любопытное свидетельство технологии хранения пива в бочках?О том, насколько бочарный промысел был тесно связан с жизнью народа, можно судить и по пословицам и поговоркам. Так вот, о недостаточном удовлетворении духовных потребностей человека говорили: «Человек не бочка, не нальешь, да не заткнешь гвоздем». Или об умирающем человеке: «Человек не бочка, по ладам не соберешь, обручами не свяжешь». В то же время, желая подчеркнуть духовную бедность человеческой натуры кого-либо, пустоту, никчемность, говорили: «В пустой бочке звону много»; «Отъелся, бочка бочкой»; «Пошел черт по бочкам» (началась безобразная попойка).
Так вот, о недостаточном удовлетворении духовных потребностей человека говорили: «Человек не бочка, не нальешь, да не заткнешь гвоздем». Или об умирающем человеке: «Человек не бочка, по ладам не соберешь, обручами не свяжешь». В то же время, желая подчеркнуть духовную бедность человеческой натуры кого-либо, пустоту, никчемность, говорили: «В пустой бочке звону много»; «Отъелся, бочка бочкой»; «Пошел черт по бочкам» (началась безобразная попойка).
В наше время бочарным промыслом, некогда процветавшим, в индивидуальной мастерской мало кто занимается, хотя спрос на бондарную посуду немалый. Да это и понятно. Бочарная продукция, разнообразная по форме и размерам, по назначению и применению и даже по художественному исполнению , находит самое широкое использование. Ею пользуются для квашения и соления, для виноделия и пивоварения, для хранения всевозможных пищевых и непищевых продуктов.
Из одной старой книги по бочарному ремеслу приведем выдержку, свидетельствующую о распространении этого дела у нас в стране в начале XX века: «Бочарное дело является в России одной из самых крупных отраслей кустарной промышленности. Трудно найти такой угол в губерниях, обладающих лесами, где крестьяне не занимались выделкой той или другой деревянной посуды. Ведется бочарство с незапамятных времен и переходит из поколения к поколению: от деда к отцу и от отца к сыну, доставляя изрядный заработок, являющийся большим подспорьем крестьянину в его хозяйстве».
Трудно найти такой угол в губерниях, обладающих лесами, где крестьяне не занимались выделкой той или другой деревянной посуды. Ведется бочарство с незапамятных времен и переходит из поколения к поколению: от деда к отцу и от отца к сыну, доставляя изрядный заработок, являющийся большим подспорьем крестьянину в его хозяйстве».
Итак, читатель уже догадался, что бочарным производством стоит заняться в том случае, если имеется лес. Но прежде чем вести речь о сырье, остановимся на некоторых общих понятиях.
Бочка и ее составные части
Из всех бондарных изделий была, есть и остается наиболее распространенной бочка, которая чаще всего бывает с выпуклым остовом. Для создания деревянной бочки используют дощечки-клепки, или лады. Из них, в свою очередь, формируют три набора. Для изготовления первого главного набора, предназначенного для боковой стенки, или остова бочки, используют изогнутые длинные и узкие дощечки-клепки. Два других набора представляют собой днища, или донья, плоской формы, большей частью округлые. Для того чтобы днища держались в ладах, в последних в обоих концах выбирают фальц, называемый уторным пазом, или просто утором. В него входят поперечные доски, составляющие донья. Сами боковые дощечки (клепки, лады) отфуговывают по боковым граням так ровно, чтобы они прилегали друг к другу очень плотно. Этому плотному прилеганию помогают стягивающие их обручи — железные или деревянные.
Для того чтобы днища держались в ладах, в последних в обоих концах выбирают фальц, называемый уторным пазом, или просто утором. В него входят поперечные доски, составляющие донья. Сами боковые дощечки (клепки, лады) отфуговывают по боковым граням так ровно, чтобы они прилегали друг к другу очень плотно. Этому плотному прилеганию помогают стягивающие их обручи — железные или деревянные.
Бочка, по В.И.Далю (от «бочковатый», «бочистый», «бок»), — вязаная обручная деревянная посудина, состоящая из ладов, или клепок, двух дон, врезанных в уторы, и обручей (рис. 1). Понятно, что свое название эта деревянная посудина получила из-за выпирающих в стороны боков. Кстати, эта конструктивная особенность бочки с выпуклым остовом (в отличие от прямого) придает ей особенную прочность. В больших бочках, если необходимо, просверливают дыру, в последнюю вставляют кран (верток) или затыкают так называемым гвоздем (затычкой).
Открытые бочарные изделия (кадушки, ведра, ушаты, чаны и т.д.) имеют одно дно. Их боковые остовы представляют собой прямые стенки, располагаемые под острым, прямым или тупым углом по отношению к плоскости дна.
Их боковые остовы представляют собой прямые стенки, располагаемые под острым, прямым или тупым углом по отношению к плоскости дна.
Размеры и объем бочек
Размеры длины клепок и днищ у бочек колеблются от 60 до 180 см. Для клепок длиной 180 см берут кряж соответствующей длины (с прибавкой 4-5 см), диаметром 40- 50 см. Из такого кряжа должно выйти 24 клепки шириной 14-16 см и толщиной 4 см.
Для клепок длиной 150 см берут кряж, имеющий в диаметре 36-40 см. Число клепок из такого кряжа получают 24, ширина каждой 10 см, толщина 4 см.
Для клепок длиной 120 см и 90 см подойдет кряж диаметром 28-36 см. Ширина клепок 8 см, толщина 3 см.
Для клепок длиной 60 см кряж берут диаметром 18-26 см. Ширина полученных клепок будет 6-8 см, а толщина 1,5-2 см.
Кряж размечают так, как показано на рис. 2,о. Затем каждую шестую часть делят на четыре. Из них уже делают клепки нужных размеров, следя за тем, чтобы заболонь и сердцевина были отколоты. В том случае, когда кряж будет больше нам необходимого для выделки соответствующего размера клепок, его можно разметить и другим способом — двухрядным или трехрядным (рис. 2,6».
2,6».
Для распиловки кряжа на клепки можно предложить следующие схемы (рис. 3,4,5,6).
Для днищ 180-сантиметровых бочек идет кряж диаметром 56-60 см, длиной 94 см. Ширина досок 30 см, толщина 3-4 см.
Чтобы изготовить 40-ведерную дубовую бочку, нужны клепки длиной 90-120 см, шириной 8-14 см, толщиной 2-3 см.
Для обычных кадок заготавливают клепки 60-90 см длиной, 8-12 см шириной. 4 см толщиной.
Для небольших бочонков и ведер изготавливают клепки длиной 60-90 см, шириной 10 см и толщиной 2-3 см.
Наиболее ходовыми бочками являются такие, у которых высота составляет 50 и 70 см. В целях более экономного расхода материалов есть смысл изготавливать бочки по парам. Одну высотой 50 см, другую высотой 70 см. В этом случае отходы более крупной бочки могут послужить заготовками для мелкой.
Из-за яйцевидной формы расчет объема бочки затруднителен. Однако на практике бондари нашли способ быстрого и довольно точного подсчета этого объема. Итак, для расчета объема бочки необходимо измерить ее высоту от одного утора до другого, а также диаметры в двух местах: в центральной части и в дне. Измерения лучше производить в дециметрах (напомним, 1 дм= = 10 см), так как 1 дм3 равен 1 л. Затем каждый измеренный диаметр возводят в квадрат.
Измерения лучше производить в дециметрах (напомним, 1 дм= = 10 см), так как 1 дм3 равен 1 л. Затем каждый измеренный диаметр возводят в квадрат.
Далее большее из полученных чисел удваивают и прибавляют к меньшему. Результат умножают на высоту бочки, а потом еще раз умножают на 3,14. Полученное от умножения произведение делят на 12 и получают объем бочки в литрах. Чтобы узнать, какое количество ведер содержится в бочке, ее объем в литрах делят на 12 (обычный объем»одного ведра в литрах).
Для примера рассчитаем объем бочки, у которой высота 70 см (7 дм), большой диаметр 60 см (6 дм), малый диаметр (поперечник днища) 50 см (5 дм). Произведем расчеты:
1) 5×5 = 25 дм2;
2) 6×6 = 36 дм2;
3) 36 х2 = 72 дм2;
4) 72 + 25 = 97 дм2;
5) 97 дм2 х7 дм = 679 дм3;
6) 679 дм3хЗ,14= = 2132 дм3;
7) 2132 дм3: 12 = 148 дм3 = = 148 л;
8) 148 л: 12 = 15 ведер.
В буквенном выражении формула расчета объема бочки будет выглядеть так:
(d2 + 2D2) h — п
где: V — емкость бочки в литрах;
d — диаметр дна бочки;
D — диаметр центральной части бочки;
h — высота бочки;
л — постоянная величина 3,14.
Какой формы и сколько нужно клепок?
Для облегчения поиска ответов на поставленные вопросы бондарь на листе картона или бумаги чертит окружности центра и дна будущей бочки (рис. 7). Причем можно чертить в масштабе 1:1. Тогда расчеты упрощаются. Или можно чертить с соответствующим уменьшением в 2, 4, 5 раз и т.д. И тогда при расчетах необходимо это уменьшение учитывать.
Итак, мы знаем, что в нашем примере большой диаметр равен 60 см. Диаметр дна 50 см. Вычерчиваем соответствующие диаметры на чертеже. Если нам известен только диаметр дна, то без особого труда (прибавив 1/5 часть диаметра дна) можно получить диаметр центральной части бочки (брюшной). И наоборот. Если нам известен большой диаметр, то можно вычислить (отняв 1/6 большого диаметра) диаметр дна.
Установить число клепок можно двумя способами. Либо, зная ширину в центре одной заданной клепки, выстраиваем на чертеже по большой окружности необходимое количество данной величины. Либо данную окружность делим на определенное число раз (в нашем случае на 16) и таким образом узнаем ширину самой широкой части клепки. Зная радиус большой окружности (30 см), по известной формуле (2tcr) находим длину этой окружности: 2x30x3,14 = 188,4 см.
Зная радиус большой окружности (30 см), по известной формуле (2tcr) находим длину этой окружности: 2x30x3,14 = 188,4 см.
Теперь эту длину делим на количество клепок (16). Получим 11,7 см. Округлив это число, получим 12 см. Такова будет ширина центральной части клепки. Если на чертеже мы проведем соответствующее число радиарных линий (в нашем случае 16), то здесь же на чертеже сможем измерить ширину конца клепки. Она составит приблизительно 10 см. То есть ширина конца клепки будет меньше ширины центральной ее части на 1/6 последнего размера.
На нашем рисунке мы можем установить также и кривизну (выпуклость) клепок и величину скоса боковых граней. Мы можем количество клепок увеличить или уменьшить. Соответственно изменятся и размеры каждой отдельно взятой клепки. Заметим, что при заданной высоте бочки 70 см от утора до утора действительная длина клепки должна быть примерно 84 см (с учетом на изгиб и на обрезку).
Толщина клепки во взятом примере будет равна 2 см (60-50 = 10 см; 10:5 = 2 см). Толще V — обший объем цилиндрического изделия; d — диаметр дна; я — постоянная величина, равная 3,14.
Толще V — обший объем цилиндрического изделия; d — диаметр дна; я — постоянная величина, равная 3,14.
Внутренний объем конических бондарных изделий вычисляют по формуле усеченного конуса:
V = л h (D2 + d2 + Dd).
Буквенные обозначения в данной формуле прежние.
Выделка клепок, или ладов
Расскажем о выделке клепок поопераци-онно.
1. Нарезка клепок. Для выделки клепок используют разные породы деревьев. В зависимости от предназначения бочек выбирают и соответствующее дерево. Скажем, лучшими бочками считаются дубовые. Их в основном предназначают для хранения спирта, коньяка, пива, вина и т.п. Для изготовления клепок под бочки, используемые в виноделии, идет обычно белый дуб.
Кстати сказать, использование дубовых бочек в виноделии очень часто является необходимым технологическим условием для получения соответствующего напитка. Так, например, спиртной напиток ром (крепостью 45%) получают из выдержанного ромового спирта, который происходит в результате сбраживания и перегонки сока сахарного тростника. Выдержка рома в дубовых бочках является непременным условием технологии.
Выдержка рома в дубовых бочках является непременным условием технологии.
Если в бочке собираются хранить воду, то для нее клепки делают из сосны, осины или ели. Для хранения молока и молочных продуктов на бочки идет можжевельник и липа.
К исходной древесине предъявляют определенные требования. Она должна быть сухой и без пороков: без отлупа, червоточины, прорости, свилеватости, исззаросших сучков, без так называемых раковин. О гнилом и прелом дереве и говорить нечего. Ясно, что таковое для выделки бочек не годится.
Для изготовления клепок лучше всего брать лес, колотый по сердцевинным слоям. Клепки из таких дощечек получаются наиболее прочными на изгиб. Обычно их вытесывают специальным бондарным топором. Но делают клепки и пилеными. Если вытесненные клепки предназначают для бочек, в которых потом собираются хранить различные жидкости, то пиленые клепки идут на бочки для сыпучих материалов — песок, мука и т.п.
Колоть клепки лучше всего из дерева, только что срубленного. А время заготовки наиболее подходящее — это октябрь и ноябрь. Деревья при помощи пилы или топора валят на землю. И тут же разделывают на клепки (рис. 10). То есть вначале дерево очищают от сучьев, затем распиливают на кряжи с таким расчетом, чтобы они по Алине на 2-3 см превышали будущие клепки или даже еще больше. Далее кряжи колят по сердцевинным лучам на части. Иногда колят и по годичным кольцам. Тогда клепка получается уже выпукло-вогнутой (рис. 11). Но по сердцевинным лучам колоть легче. Удобно колоть топором-колуном, у которого обух толстый, а клин острый и широкий.
А время заготовки наиболее подходящее — это октябрь и ноябрь. Деревья при помощи пилы или топора валят на землю. И тут же разделывают на клепки (рис. 10). То есть вначале дерево очищают от сучьев, затем распиливают на кряжи с таким расчетом, чтобы они по Алине на 2-3 см превышали будущие клепки или даже еще больше. Далее кряжи колят по сердцевинным лучам на части. Иногда колят и по годичным кольцам. Тогда клепка получается уже выпукло-вогнутой (рис. 11). Но по сердцевинным лучам колоть легче. Удобно колоть топором-колуном, у которого обух толстый, а клин острый и широкий.
Из рисунка 10 видно, как производят эту работу и в какой последовательности. В зависимости от толщины кряж колят вначале на половины, затем на четвертины, на восьмые доли. Если возможно, то колят и на шестнадцатые доли и т.д. От полученной минимальной части кряжа откалывают заболонь и сердцевину — то есть самые рыхлые слои древесины вместе с корой при помощи клиновидного изогнутого ножа (см. рис. 11). Теперь полученную среднюю часть колят по годичным кольцам надвое или натрое. Вновь полученные части называют гнатин-ником. По ширине его стараются получить на 1 см больше ширины будущей клепки (рис. 12). А вот уже гнатинник разделывают на клепки. Ясно, что и по толщине заготовки должны превышать толшину будущей клепки: ведь сырой лес, высыхая, на 12- 20% сократится. Бондарь по опыту знает, каких размеров ему делать заготовки в зависимости от породы и влажности леса.
Вновь полученные части называют гнатин-ником. По ширине его стараются получить на 1 см больше ширины будущей клепки (рис. 12). А вот уже гнатинник разделывают на клепки. Ясно, что и по толщине заготовки должны превышать толшину будущей клепки: ведь сырой лес, высыхая, на 12- 20% сократится. Бондарь по опыту знает, каких размеров ему делать заготовки в зависимости от породы и влажности леса.
Мы уже видели схемы однорядной, двухрядной и трехрядной выколки кряжей. Заметим, что больше всего отходов получается при однорядной выколке. Это наглядно видно на рис. 13 при сравнении его с рис. 2,б,в.
Сухое дерево колется труднее. Пилить клепки, естественно, из сухого дерева легче. Клепки нарезают с таким расчетом, чтобы в середине они были шире, чем по концам (точнее, их потом стесывают). Но по концам их толщина несколько больше, чем в средней части. Утолщение по концам необходимо для вырезки потом утора, то есть паза под дно или днище. Для правильной и более быстрой нарезки клепок используют шаблон. В качестве последнего может послужить уже готовая клепка. Можно также изготовить шаблон из фанеры в виде готовой клепки.
В качестве последнего может послужить уже готовая клепка. Можно также изготовить шаблон из фанеры в виде готовой клепки.
2. Сушка клепок . Прежде чем окончательно отделывать клепки, их сушат. Клепки складывают по две крест-накрест. Естественная сушка может длиться до года. Поэтому обычно бондарь делает себе запас клепок на это время. Сушить клепки можно и в специальной сушилке — закрытом помещении с подогревом и с обеспечением циркуляции воздуха.
Если бондарь делает бочки, как говорится, для своей надобности, то необходимость в устройстве специальной сушильни отпадает. Ведь для изготовления одной-двух бочек клепки можно высушить и дома над печкой или без нее, если дом не сельский и не дачный. При сушке следят за тем, чтобы клепки не растрескались, особенно на торцах. Для этого последние смазывают глиной или краской или еще заклеивают бумагой. По времени сушка может длиться от одного дня (например, на жарко истопленной печи) до нескольких дней (в теплом помещении).
3. Обработка клепок. После сушки доски как клепок, так и днищ обрабатывают, то есть им придают именно ту форму, которая необходима для изготовления бочек.
Обработка клепок. После сушки доски как клепок, так и днищ обрабатывают, то есть им придают именно ту форму, которая необходима для изготовления бочек.
Обычно клепки делают на 2-3 см длиннее, чем необходимо, поэтому после сушки их укорачивают с обоих концов лучковой пилой. Если бочку делают с вогнутым дном, то клепки не укорачивают, а спиливают, выравнивают в седле, когда бочка собрана, связана обручами и уже намечено место для дна.
Просушенные и укороченные клепки обрабатывают внутри и снаружи. Обрабатывает их каждый бондарь по-своему. В результате обработки клепки должны быть очень точно подогнаны друг к другу.
Вначале обработки клепку стесывают с наружной стороны специальным бондарным топором (он сточен на одну сторону). Работает бондарь на чурбане (рис. 15), держа клепку левой рукой и производя стесывание правой. Можно стесывать не только топором, но также одним из стругов или косарей на бондарной скамье (рис. 16, 17). Движения бондаря при этой работе должны быть неторопливыми, очень расчетливыми, с тем чтобы не испортить клепку излишним отщепом или надрубом. Как правило, косарями (рис. 18), шляхтами (рис. 19) и стругами (рис. 20) бондарь пользуется для последующей чистовой обработки клепки. Обтесанную снаружи и внутри клепку сверяют с шаблоном. Когда обтесывание закончено приступают к обстругиванию клепки. Для этой цели сначала берут рубанок с выпуклой подошвой и с лезвием дугообразной формы Им обстругивают клепки, а затем слегка сглаживают последние прямым рубанком, снимая небольшую стружку. Окончательную отделку и обработку клепок осуществляют тогда, когда они уже собраны в бочку. На рис. 21,с показана клепка такой формы, какой необходима для изготовления выпуклых бочек. Форма может быть и такой, какой она показана на рис. 21,6″, Эта клепка в середине гораздо шире, чем по краям. Скашивают клепку к краям очень аккуратно. Работу эту можно делать на глаз, но лучше, все время сверяя с шаблоном, отмечая неровности карандашом. В выполнении данной работы нужна не только аккуратность, но еще и большая точность. Если ее не будет, то при сборке бока клепок могут не сойтись, и хлопот тогда по пригонке не оберешься.
Как правило, косарями (рис. 18), шляхтами (рис. 19) и стругами (рис. 20) бондарь пользуется для последующей чистовой обработки клепки. Обтесанную снаружи и внутри клепку сверяют с шаблоном. Когда обтесывание закончено приступают к обстругиванию клепки. Для этой цели сначала берут рубанок с выпуклой подошвой и с лезвием дугообразной формы Им обстругивают клепки, а затем слегка сглаживают последние прямым рубанком, снимая небольшую стружку. Окончательную отделку и обработку клепок осуществляют тогда, когда они уже собраны в бочку. На рис. 21,с показана клепка такой формы, какой необходима для изготовления выпуклых бочек. Форма может быть и такой, какой она показана на рис. 21,6″, Эта клепка в середине гораздо шире, чем по краям. Скашивают клепку к краям очень аккуратно. Работу эту можно делать на глаз, но лучше, все время сверяя с шаблоном, отмечая неровности карандашом. В выполнении данной работы нужна не только аккуратность, но еще и большая точность. Если ее не будет, то при сборке бока клепок могут не сойтись, и хлопот тогда по пригонке не оберешься.
О внутренней обработке клепки скажем несколько подробнее. При этой работе прежде всего намечают толщину клепки по всей поверхности, особенно старательно в шейках, то есть на концах. Толщину отмечают при помощи шаблона — чертилки (рис. 22). Чертилку накладывают в середине клепки так, чтобы острие а пришлось на самое ребро клепки. Затем шаблон ведут по всей длине клепки. Острие б отметит толщину шейки. Понятно, что при изготовлении бочек разных размеров разными будут и толщины клепок. А следовательно, и чертилок у бондаря должно быть несколько. Клепку с размеченной толщиной укрепляют в станке и всю лишнюю древесину стесывают топором или стругом.
Последняя операция по обработке клепок — это их прифуговка. Как мы уже говорили, очертания будущей бочки напрямую связаны с формой клепки. Если у клепки боковые линии будут прямыми, то и бочка получится прямой. Самая прочная и удобная форма бочки выпуклая. Для нее клепку делают такой, какой она показана на рис. 21. То есть у нее середина широкая, концы сужены. Наиболее распространенное соотношение середины и концов клепки, как мы уже отмечали, следующее: на конце клепка должна быть уже или меньше середины на 1/6 часть. К примеру, если в середине ширина клепки 12 см, то в концах она будет равна 10 см. Соотношение может быть и другим. Заметим, чем больше разница между шириной в середине и в конце клепки, тем круче будет бочка в боках.
Наиболее распространенное соотношение середины и концов клепки, как мы уже отмечали, следующее: на конце клепка должна быть уже или меньше середины на 1/6 часть. К примеру, если в середине ширина клепки 12 см, то в концах она будет равна 10 см. Соотношение может быть и другим. Заметим, чем больше разница между шириной в середине и в конце клепки, тем круче будет бочка в боках.
Обстругивают и прифуговывают размеченные ребра клепки рубанком и фуганком, закрепляя ее в ладиле (рис. 23). А можно эту операцию выполнять и на большом бочарном рубанке (рис. 24). При фуговании ребра не прифуговывают вплотную, а делают небольшой зазор. То есть ребра клепок чуть скашивают внутрь. При стягивании бочки обручами имеющийся зазор исчезнет: клепки плотно прижмутся друг к другу.
Днища
Эти части бочки изготавливают из досок, которые чуть толще клепок. Доски обстругивают сначала рубанком, а затем прифуговывают плотно друг к другу. В зависимости от ширины досок и размера бочки днище может быть сколочено из четырех, пяти, шести и т. д. досок (рис. 25). Удобнее дощечки для днища нарезать из одной доски. Так как днише у бочки имеет круглую форму, то и составные дощечки подбирают такой длины, чтобы потом при придании днищу округлости было меньше отходов (рис. 26). Строгают доски днища, как правило, снаружи. Изнутри либо совсем не строгают, либо строгают только слегка.
д. досок (рис. 25). Удобнее дощечки для днища нарезать из одной доски. Так как днише у бочки имеет круглую форму, то и составные дощечки подбирают такой длины, чтобы потом при придании днищу округлости было меньше отходов (рис. 26). Строгают доски днища, как правило, снаружи. Изнутри либо совсем не строгают, либо строгают только слегка.
Обручи
Их делают либо железными, либо деревянными. Железные изготавливают из полосного железа, ширина которого зависит от величины бочки . Чаще всего ширина составляет 3-4 см. Концы полосного железа накладывают друг на друга и склепывают. Железные обручи целесообразно применять для крупных бочек. Для деревянных обручей используют древесину клена, дуба, вяза, бука, ясеня. Применяют для деревянных обручей и какое-либо другое прочное и гибкое дерево — можжевельник, черемуху, ель и т.д. Для обручей выбирают молодое дерево, которое подрезают каждые 10-12 лет, — оно наиболее гибкое. При заготовке древесины для обручей пользуются следующими инструментами: топор, нож, струг, мялка, щепальные клинья, или колонки. Хорошо заготавливать деревянные обручи поздней осенью или в начале зимы. Кору с молодых деревцев или прутьев не снимают. В зависимости от толщины каждый прут расщепляют вдоль на две половины, на три или на четыре части.
Хорошо заготавливать деревянные обручи поздней осенью или в начале зимы. Кору с молодых деревцев или прутьев не снимают. В зависимости от толщины каждый прут расщепляют вдоль на две половины, на три или на четыре части.
Чтобы расщепить на две пластины, удобно пользоваться ножом. В остальных случаях пользуются щепальным клином, изготовленным из твердых пород дерева (рис. 27). В пруту делают надрез ножом на три или четыре части. Вставляют в надрез соответствующий щепальный клин и натягивают на него прут. Последний расщепляется на нужное нам количество частей. Чаще всего обручи делают из половинок прута, которые сгибают вокруг вбитых в землю по кольцу кольев (рис. 28). Концы обручей заводят за колья. Зафиксировав таким образом обручи, им дают высохнуть. Но удобнее для гнутья обручей пользоваться специальной конусообразной болванкой (рис. 29). Верхняя часть этой болванки соответствует малым обручам, нижняя — большим. Иногда заготовки, перед тем как согнуть в обручи, распаривают. Для удобства гнутья пользуются вспомогательными инструментами — мялкой или специальной скобой вбитой в стену либо в деревянный брус (рис. 30).
Сборка клепок
После того как подготовлены клепки, днища, обручи, приступают к сборке бочки. В первую очередь, конечно, собирают клепки. Но, перед тем как собирать их, клепки необходимо, по выражению бондарей, причертить друг к другу, то есть подогнать, прижать. Причерчивают при помощи обычного циркуля, рейсмуса или штангенциркуля. Отыскивают на концах каждой клепки середину и отмечают ее. Далее находят середину по длине клепки и, поставив здесь острие неподвижной ножки циркуля, другим концом очерчивают дугу на концах клепки. Проделав эту операцию со всеми клепками, находят таким образом шейную линию. Именно по ней потом придутся уторы для вставки днищ.
После расчерчивания приступают к сборке клепок. Берут вначале головной, или торцевой обруч (тот, которым стягивают клепки по концам) и прикрепляют к нему втулочную клепку. Таковой называют ту клепку, в которой будет находиться втулка бочки, если она запланирована. Втулочную или обычную первую клепку прикрепляют к обручу при помощи струбцинки или зажима, похожего на бельевую прищепку (рис. 31).
Оговоримся, в бондарных мастерских начинают сбор остова бочки при помощи специального рабочего обруча. Он представляет собой металлическое кольцо из круглого или полосового железа толщиной 10- 15 мм. Диаметр рабочего обруча обычно несколько больше диаметра постоянного — ведь его потом убирают, заменяя последним. В зависимости от размеров бочки в бондарных мастерских имеется несколько рабочих обручей, которые дублируют постоянные (головные, они же шейные или торцевые, срединные, или брюшные). Используют также и предохранительный обруч, который, в сущности, тот же рабочий (рис. 32).
Итак, продолжим речь о сборке клепок в остов. Прямо напротив первой клепки ставят самую широкую или основную клепку, а между ними по бокам на одинаковом расстоянии еще по одной. Клепки также закрепляют струбцинками или зажимами. Подобная расстановка клепок поможет прочно удержать головной обруч как бы на четырех ножках. Далее остальные клепки расставляют на свои места. Затем снимают зажимы и головной обруч несколько осаживают книзу, одновременно нагоняя на остов один или два шейных обруча и один средний (его еще называют брюшным, или пуковым). Можно эту первоначальную работу по сбору клепок в остов проделать иначе. То есть, поставив две клепки напротив друг друга, накладывают обруч и устанавливают другие клепки одну за одной, прикрепляя зажимами. Конечно, трудно приготовить клепки , которые подошли бы друг к другу, как говорится, без сучка и задоринки.
Случается так, что последняя клепка оказывается более широкой, чем необходимо. Тогда одну или две соседние клепки уменьшают в ширине. Или одну широкую заменяют двумя узкими клепками. В том случае, если не совпадают диаметры краев бочки, то есть один край шире или уже другого, две-три или несколько клепок перемещают концами в обратную сторону. Таким образом добиваются равенства диаметров у верхнего и нижнего основания бочки. Когда все клепки расставлены, надеты шейный и срединный обручи, остов переворачивают и стягивают клепки при помощи ворота (рис. 34) или веревки (рис. 35). Однако стягивать клепки нужно осторожно, чтобы не поломать какие-либо из них. Лучше всего стягивать предварительно распаренные клепки. Есть несколько способов нагрева и распаривания последних. В больших бочарных мастерских пользуются специально устроенной мангальной печью с жаровым колпаком (рис. 36). Принцип ее работы понятен из рисунка. Для мастерских меньшего размера можно порекомендовать железный таганчик-мангалку (рис. 37). Производят распарку клепок при помощи еще железной круглой печи с надставной трубой.
Дупло (так бондари называют полусобранный остов) надевают на эту печь. Ее топят, а клепки с внутренней стороны предварительно смачивают водой. При нагревании происходит распаривание клепок. После этого они становятся более податливыми на изгиб, менее ломкими. Если диаметр бочки меньше нашей круглой печки, то дупло надевают на печную трубу, предварительно сняв у нее одно колено, а потом (после размещения дупла), поставив его на место. Теперь печная труба, проходящая через дупло бочки, будет производить нужную нам по распариванию работу. Само дупло размещают на подставах, сверху и снизу прикрывая железными крышками. Каждую из крышек вырезают из листового железа в виде двух полукругов с аналогичными полукруглыми вырезами для пропуска печной трубы. Опять обильно обрызгивают дупло водою перед распаркой, да и во время оной. Вода от жара печной трубы нагревается, превращаясь в пар. Ну а последний делает свое распаривающее дело. Сколько распаривать клепки — каждый бондарь решает опытным путем. Обычно эта операция длится 1-2 часа. Слишком распаренные клепки становятся чересчур мягки на изгиб. Недораспаренные клепки при сгибе лопаются.
Продолжительность распаривания зависит также и оттого, насколько клепки необходимо согнуть. Если мы изготавливаем небольшую бочку с малым изгибом клепок, то необязательно прибегать к помощи железной круглой печи. Можно воспользоваться и железным таган-чиком-мангалкой. В мангалке разжигают дрова. Когда образуются жарко тлеющие угли, ее ставят в середину дупла и производят распаривание клепок. Разумеется, эту работу проделывают в каком-либо нежилом помещении, где существует свободный обмен с наружным воздухом. Распаренные клепки стягивают. Делают это, как уже отмечали, при помощи затяжек и ворота или при помощи обычной палки и веревки (закрутка). На шейную часть остова набрасывают веревочную петлю и постепенно ее затягивают. Если имеющиеся клепки толстые (как правило, у больших бочек), то используют не одну, а две, а то и три затяжки. Затягивают постепенно. Сначала стягивают срединную часть, затем шейную. Полезно дупло бочки крутонуть то в одну, то в другую сторону, поворачивая как рулевое колесо у автомобиля. Это помогает стяжку клепок сделать равномерной. Порою то одна, то другая клепка выпирает из общего ряда. Ее вправляют при помощи деревянного молотка — киянки. Когда концы клепок достаточно плотно сойдутся, на дупло бочки нагонять начинают обручи. Вначале большой (брюшной), затем шейный и головной. Эти обручи рассматриваются как рабочие. Постоянные обручи нагоняют на бочку после вставки днищ.
После того как клепки стянуты с одной стороны дупла, его переворачивают и дотягивают клепки на другом конце. Полученный предмет со стянутыми клепками уже по праву называется остовом бочки, или бездонной бочкой. Этот остов с рабочими обручами сушат в течение нескольких дней или одной-двух недель (в зависимости от условий сушки: около печки или на открытом воздухе). Затем его закаливают изнутри, то есть подвергают обжигу. Для этого в остове разжигают стружки. Далее остов перекатывают, следя за тем, чтобы древесина не обуглилась, а лишь слегка подкалилась, приобретя золотистый оттенок. Так поступали старые мастера. Но проще подвергнуть остов опаливанию паяльной лампой, соблюдая, естественно, правила противопожарной безопасности. Обжиг или закаливание производят дня того, чтобы клепки в остове стали значительно устойчивыми по форме. В промышленных условиях закаливание производят на мангааьной печи. Небольшие бочонки можно и не обжигать. Их достаточно высушить при высокой температуре, например, в духовке русской печи.
Остовы конической формы (с прямыми стенками) вообще не закаливают, так как их клепки не имеют изгиба по длине. После закалки бездонной бочки ее обручи осаживают, так как во время обжига древесина размягчилась, часть ее влаги испарилась, то есть клепки несколько усохли. Осаживают обручи при помощи молотка и набойки (рис. 38, 39, 40). Во время этой операции клепки своими ребрами плотно прижимаются друг к другу, не оставляя никаких щелей или зазоров. Все неровности просто-напросто сминаются. Затем приступают к обрезке выступающих концов клепок лучковой пилой, расположив остов в седле (рис. 41) или на скамье (рис. 42).
Как производят это выравнивание, видно из последнего рисунка. Заметим только, что обрезку ведут так, чтобы поверхность пропила была на клонена несколько внутрь остова. Далее снимают фаски при помощи бондарного ножа, струга или бочарного рубанка. Съему фасок или срезов производят на половину толщины торцов. Таким образом, предупреждают какие-либо откалывания концов клепок, их расщепливание на внутренней стороне остова. Торцы последнего после съемки фасок вообще приобретают опрятный и красивый вид. Здесь мы лишний раз убеждаемся в том, что красота и польза не разделимы, они очень тесно взаимосвязаны.
Снаружи края торцов мы пока не трогаем. Их отделку оставляем на потом, когда завершим изготовление бочки. Перед тем как вырезать уторы и вставлять днища, остов бочки остругивают изнутри и снаружи. Дело в том, что после обжига и осаживания обручей кромки соседних клепок часто образуют выступы (бондари называют их провесами). Вот эти провесы и необходимо сгладить при помощи стругов . Для наружного остругивания применяют вогнутый струг, скобель или рубанок, для внутреннего — выпуклый.
При остругивании снаружи временно снимают поочередно обручи. Сначала с одного конца остова, затем с другого. Особенно тщательно выравнивают шейную поверхность остова изнутри. Только в этом случае удается утор-ный паз выбрать ровным и по окружности и по глубине. А следовательно, и вставка днищ будет плотной и прочной. Иногда этой зачисткой шейной части на расстоянии 10- 15 см от края остова и ограничиваются.
После окончания зачистки приступают к выемке уторного паза. Эту операцию производят уторником (рис. 43). А если бочарное изделие небольшое и чистота, и правильность выемки не требуются, то уторный паз выбирают гребнем (рис. 44). И в том и в другом случае от края отступают 3-5 см.
Уторный паз выбирают только с одной стороны, если готовят бочку, открывающуюся с другого конца. Если предполагают изготовить бочку глухую, двухдонную (закрытую), то уторный паз выбирают в двух концах остова. Для совершения этой операции остов бочки размещают в седле или на верстаке. При выемке уторного паза бондари пользуются несложным правилом. Глубина паза не должна быть больше половины толщины концов клепок, а ширина утора не должна превышать толщину досок днища. Даже наоборот — ширину делают несколько уже толшины днища примерно на 3-5 мм. Так только можно будет добиться плотности прилегания днища в бочке и предотвратить возможную течь.
Теперь приступаем к изготовлению днищ. Хотя об этом речь уже шла выше, напомним, что днища делают из клепок-дощечек, разных по ширине, но одинаковых по толщине, плотно подогнанных и прифугованных друг к другу. Толщина днищ обычно превосходит толщину боковых клепок. В зависимости от величины бочарного изделия днища могут состоять из 4-6 дощечек, сплоченных в один щит. Прежде чем сплачивать дощечки в единый щит, каждую из них тщательно строгают стругом, скобелем, рубанком.
Также тщательно, а может, даже и более тщательно отфутовывают боковые грани. После этого дощечки зажимают в щемиле (рис. 32). Можно предварительно их сплотить при помощи шипов. На образовавшемся из дощечек щите, зажатом в щемиле, очерчивают круг будущего днища (рис. 26). Внимание — его диаметр должен превосходить диаметр бочки в уторе на двойную глубину уторного паза.
Теперь лишние части досок отпиливают лучковой пилой по сделанной разметке. Можно предварительно щит разобрать. А можно производить опиловку его прямо в щемиле. Внешнюю сторону днища еще раз тщательно остругивают. С внутренней стороны у днища стесывают края. Циркулем очерчивают границу этой отлогой фаски. Ее ширина обычно составляет 4-7 см.
Снимать эту фаску необходимо потому, что толщина досок днища больше толщины угарного паза. При снятой фаске днище зайдет в утор и по мере захода будет возрастать плотность соприкосновения его с угарным пазом. Иногда фаску снимают и с наружной стороны днища. Но фаску эту делают небольшой. По своей ширине она должна быть меньше глубины уторного паза. Тогда после вставки дна в бочку фаска полностью спрячется.
Доски, составляющие днище , каждая имеет свое название. В днище, состоящем из 4 досок, две срединные называются главными, а боковые — отрезами. В днище из 6 досок две средние также называются главными, следующие две — боковыми, а крайние — по-прежнему отрезами. Подготовленное днище вставляют в утор. Целиком днище вставлять трудно. Чаще его вставляют разобранными дощечками. Предварительно с конца остова бочки снимают один или два обруча.
Клепки при этом разойдутся. Вставляют днище, начиная с крайних (боковых) дощечек. Последнюю среднюю дощечку вставить труднее всего. Вставляют ее примерно в такой последовательности. Вначале заводят один конец в уторный паз. На другом краю отгибают одну или две клепки так, чтобы сподручно можно было завести и второй конец дощечки в утор. Совершая эту работу, пользуются вспомогательным инструментом: накидными клещами (рис. 32), натягом (рис. 45). Клепки при вставленном днище несколько разойдутся.
Их вправляют на место деревянным молотком. Вставив днище на одном конце бочки, вставляют аналогично и на другом. Второе дно вставить труднее, так как его уже нельзя поддержать снизу.
Не по одной дощечке, а целиком днище вставляют в следующем порядке. Сначала вводят в утор один торцевой край. Далее разводят широко клепки и вводят в утор днище целиком. Перед вставкой часто уторы промазывают с помощью лопаточки замазкой (смесь сурика или мела и вареного льняного масла — олифы). Для более плотного прилегания днища используют еще так называемую бочарную траву: ситник, тростник и т.п. Эту бочарную траву в уторном пазу размещают при помощи конопатки (рис. 38). После того как оба днища введены в уторы, клепки еще раз подправляют деревянным молотком, а потом их плотно стягивают с помощью затяжек. Завершают работу тем, что на концы бочки вновь надевают обручи.
Иногда для большей прочности днище бочки укрепляют наладной дощечкой (рис. 46) — набойкой. Она представляет собой дощечку 15 см в ширину и 3-4 см в толщину. Длина ее соответствует диаметру днища. Набойку закрепляют поперек досок днища нагелями. Последние вколачивают в концы клепок рядом с уторным пазом. Нагели делают достаточно длинными с тем, чтобы крепление набойки было надежным. Форма нагелей необязательно должна быть круглой. Онаможетбыть граненной, например четырехугольной. Даже лучше, если она будеттаковой, так как при высыхании бочки круглые нагели порою выпадают, а граненные задерживаются. Число нагелей с каждой стороны набойки варьируется от 4 до 6.
Последняя завершающая операция по изготовлению бочек — набивка постоянных обручей. Число их различно. На большую бочку набивают до 18 деревянных обручей или 6-8 железных. Для бочки средних размеров обычное число деревянных обручей — 14-16 штук. Их градация следующая: 8 шейных (по 4 обруча с каждого края), 6 брюшных (по 3 обруча в половине бочки). Реже насаживают 10 деревянных обручей (6 шейных, 4 брюшных; и шейные и брюшные обручи поровну распределяются на обеих половинах бочки). Заметим сразу же, что бочка с 10 деревянными обручами менее крепкая, чем с 14.
Деревянные обручи изготавливают из обручных хлыстов. Этими хлыстами опоясывают бочку в том месте, где предполагают разместить обруч. Делают на хлысте и на бочке соответствующие пометки. На хлысте отмечают места зарубок для вязки замка (рис. 47). На замок оставляют припуск в 10-12 см с обоих концов обруча. Сами концы срезают наискось в виде заостренных языков. Там, где отмечены у нас были зарубки, делают надрезы на половину ширины обручного хлыста. На одном конце обруча надрез делают сверху, на другом — снизу. С внутренней стороны обруча, по направлению от надрезов к середине, делают выемки длиной 4-5 см, постепенно сходящие на нет. Теперь вяжут замок. А именно: концы обруча зацепляют друг за друга выступами надрезов, укладывают в соответствующие выемки. То есть концы заводят и прячут на внутренней стороне обруча. Часто обруч в месте вязки замка для прочности оплетают ивовыми прутьями.
С бочки, как уже понял читатель, рабочие обручи снимают, заменяя их постоянными. Делать это нужно последовательно: сначала заменяют брюшные обручи в одной половине бочки, затем шейные все в той же половине, а уж потом тоже самое совершают со второй половиной бочки. Последние шейные обручи особенно трудно натягивать на остов бочки. Обруч заводят на клепки сначала с одного края.
Затем с другого, помогая при этом себе натягом и затяжками. Натягом работают так. Конец его ручки упирают в бок бочки, на другой конец все той же ручки надавливают рукой. Обруч в это время захватом натяга слегка растягивается и, охватив концы клепок, стягивает их. Клепки постепенно одну за другой глубже заводят в обруч.
Иногда надетая полуокружность обручасоскальзываете клепок. Чтобы этого не происходило, надетую половину обруча закрепляют на края остова небольшими гвоздями. Вбивать их следует не более чем на половину толщины концов клепок. После того как деревянный обруч натянут на бочку, его необходимо осадить на предназначаемое место.
Пользуются при этом деревянным молотком и набойкой (рис. 48). Набойку ставят углублением подошвы на ребро обруча. Ударами молотка по головке набойки обруч осаживают на место. Последний должен быть набит на бочку без каких-либо перекосов, до отказа, плотно охватывая» ее окружность.
Изготовление железных обручей сходно с изготовлением деревянных. Ширина и толщина железных обручей зависят от размеров бочки. Обычно берут полосное железо 3-4 см шириной. Здесь также начинают работу с обмера бочки. Отрезают полосное железо с припусками с обоих концов обруча на накладку в 10-12 см. Ножницами или зубилом отсекают также уголки концов обруча. Затем эти концы либо сваривают, либо склепывают. Сварку можно производить так, как делали бондари в старину без сварочного аппарата.
В кузнице концы обруча раскаляли докрасна. А потом, не давая остыть, на наковальне, держа щипцами и ударяя кузнечным молотом, концы сваривали. Но чаще все же концы склепывают. Их накладывают друг на друга и высверливают либо пробивают минимум две дырки, отступая от края по длине обруча 2 и 6 см. Поверхности обруча при помощи молотка придают уклон с тем, чтобы он плотнее облегал окружность бочки.
Насаживают железные обручи аналогичным способом, как и деревянные. Только при этом пользуются уже железными и молотком и набойкой. Чтобы предупредить ржавчину, железные обручи окрашивают черной масляной краской. Вид у готового деревянного изделия с черными полосами обручей — загляденье.
После насадки постоянных обручей бочку отделывают окончательно. Проходят стругом или шлифчиком по днищам и бокам бочки. Обрезают концы бочарной травы около уторов, подчищают вылезшую из них замазку. Скобелем подправляют фаски. Если запланировано, то в бочке высверливают втулочное отверстие. Стенки отверстия делают либо отвесными, либо под наклоном.
По материалам журнала: САМ
Одной из интереснейших идей, которую можно положить в основу успешно развивающегося бизнеса, является производство бочек. Они нашли свое применение еще в давние времена, когда исполняли роль сосуда для хранения вина и воды. Кроме того, они также использовались как ванны для купания.
В настоящее время деревянные бочки не являются столь популярными в использовании, как это было ранее. Объяснение этому довольно простое – дело в том, что процесс производства этих изделий существенно упростился после появления дешёвых современных технологий и материалов.
Как результат, сейчас бочки изготавливают даже из металла и разнообразных видов пластмасс. Как бы там ни было, производство бочек полностью не исчезло. Более того, современные мастера способны создать точные копии экземпляров, которые производились еще нашими предками. Как показывает практика, на этом можно также хорошо заработать.
Бочки для спиртных напитков
Одной из самых распространённых сфер применения деревянных бочек в наше время является хранение алкогольных напитков. Чаще всего ими выступают вино, коньяк и виски. В данном случае самым удачным материалом для них считается дуб, поскольку это дерево позволяет сберечь аромат и вкус напитка. Именно в таких ёмкостях хранятся элитные и дорогие вина. В связи с этим производство дубовых бочек как вид предпринимательской деятельности выглядит весьма перспективно.
Бочки для солений
Засолы – это вторая по популярности сфера применения данных сосудов. В них отлично сберегается квашеная капуста, соленые помидоры и огурцы. Более того, в таких ёмкостях очень удобно вымачивать мясо перед его копчением либо вялением.
В данном случае можно использовать как бочонки небольшого размера, так и более внушительные сосуды, в которые помещается большое количество продуктов. В любом случае незабываемый их вкус достигается именно за счёт использования деревянных бочек, а не стеклянных банок.
Бочки в дизайне
Производство бочек для декорирования интерьеров помещений является сравнительно новой идеей. Многие современные дизайнеры, которые мыслят креативно, устанавливают такие ёмкости даже вместо хорошо знакомой привычной ванны.
Такая задумка как будто переносит человека в прошлое, ведь именно бочки много лет назад использовались в виде ёмкости для купания. Особенно такая тара востребована в коттеджах, расположенных за городом. Как бы там ни было, дубовая бочка, размещённая на приусадебном участке, в гостиной, кухне либо другой комнате, выглядит необычно и оригинально.
Базовые знания в бондарном ремесле
Для того чтобы процесс действительно приносил хорошую прибыль, прежде всего это занятие должно нравиться. И в нём необходимо разбираться. Для чего, в свою очередь, рекомендуется получить определённые навыки в бондарном ремесле. Не стоит жалеть для этого времени, поскольку отсутствие понимания тонкостей, с которыми связано изготовление бочек из дуба, не приведёт к желаемому результату.
Получить их можно, выучившись у профессионального бондаря либо устроившись на некоторое время на работу на предприятие, что специализируется на данной сфере деятельности. Как показывает практика, для того чтобы досконально изучить данный процесс, необходимо самостоятельно собрать хотя бы несколько десятков деревянных бочек.
Открытие мастерской
Для того чтобы заниматься изготовлением деревянных бочек как бизнесом, изначально необходимо зарегистрировать свою фирму. В данном случае можно получить статус юридического лица либо частного предпринимателя. Кроме этого, необходимо стать на учёт в органах налогообложения.
Для открытия своего предприятия, специализацией которого будет производство бочек, требуется помещение, размер которого зависит от планируемых объёмов производства, склад для хранения готовой продукции, а также материал. В роли последнего выступает дерево и металлические обручи, которые необходимы для скрепления бочек. Помимо всего прочего понадобится персонал.
При организации такого вида предпринимательской деятельности самым сложным считается поиск квалифицированных специалистов, которые будут выполнять заказы на предприятии. Как показывает практика, в отдельных городах зачастую бывают случаи, когда их найти или вообще невозможно, или мастера из соседних населённых пунктов попросту не соглашаются переезжать. Выход из данной ситуации только один – обучить своих работников самостоятельно.
С чего начинается производство
Технология изготовления дубовых бочек является достаточно простой. На начальном этапе производится заготовка клепки, которая представляет собой небольшие дощечки, что имеют прямоугольную форму и особо выраженную фактуру. В роли сырья для них чаще всего выступает дуб.
Вместе с этим нельзя не отметить тот факт, что, в зависимости от сферы применения, зачастую используются и другие породы дерева. Исходя из технологии, каждую из клепок необходимо на протяжении как минимум двух лет просушить под открытым навесом. Благодаря этому бочка получится долговечной и высококачественной.
Сборка бочки
На втором этапе производится непосредственная сборка ёмкости. Какое-то особое оборудование для производства бочек из дерева не применяется – как и двести лет тому назад их собирают вручную. Исходя из технологии, предварительно необходимо отправить на некоторое время клепку в специально предназначенную для этого парилку. После того как древесина увлажнится и прогреется в ней, она станет более гибкой. Благодаря этому, нивелируется риск ее поломки во время сборки.
Завершающий этап производства
Последний этап изготовления деревянных бочек представляет собой сборку остова. В данном случае происходит стыковка дощечек, в результате которой изделие получает свою форму (начитываются десятки ее вариантов). После того, как крышка и дно будут выставлены, конструкция закрепляется путём использования металлических либо деревянных обручей. Далее бочка обжигается на протяжении примерно 30 минут на огне, после чего полируется и вскрывается специально предназначенными для этого составами.
Выводы
На словах процесс производства деревянных бочек выглядит немного проще, чем на самом деле. Ремеслу этому обучиться не так уж и сложно – для этого нужно только желание. Такая продукция в настоящее время на рынке довольно востребована. При этом профессионального мастера найти не так уж и просто. В связи с этим не стоит затягивать с открытием бизнеса, ведь, заняв пока что свободную нишу на рынке, можно хорошо заработать.
Бочки для солений – как их подготавливать и использовать
Чем отличается засолка в бочке или кадке от закрутки в банки?
Деревянные емкости используются в хозяйстве и, в частности, на кухне с давних времен. Сегодня искусство бондаря уже перестало быть кустарным, хотя отдельные мастера, изготавливающие дубовые бочки, кадушки и шайки все еще встречаются в сельской местности. В целом же изготовление этих необходимых для консервации емкостей поставлено на промышленный уровень, хотя объемы такой продукции не особо велики из-за малого спроса.
Деревянные емкости для солений
Однако пока есть необходимость заготавливать засолкой и квашением овощи, будут и кадки. Чем же деревянные дубовые бочки лучше металлических оцинкованных или пластиковых, которые значительно дешевле? Начать стоит с того, что дуб, из которого обычно и делают бочонки под засолку, имеет в структуре древесины дубящие вещества, что можно понять даже по названию породы дерева. Именно они являются лучшими природными консервантами, благодаря чему долго хранятся в рассоле огурцы, помидоры, капуста, грибы или рыба.
В емкости из дубовых досок быстро погибают микробы, вызывающие гниение, поскольку создается неблагоприятная для них среда. Кроме того, деревянные бочки придают новые ароматы и привкусы хранящимся продуктам, полностью изменяя в лучшую сторону вкус консервации. В этом преимущество древесины перед металлом или пластиком, которые могут несколько ухудшить вкус заготовок.
Как подготовить бочонок из дерева к хранению продуктов?
Первое, что следует помнить – если прошлогодняя консервация съедена, ни в коем случае нельзя оставлять бочонок до следующего урожая с остатками рассола или, тем более, пустым, пусть даже и вымытым. Прежде всего, нужно полностью удалить следы закончившейся заготовки, будь то соленые огурцы или квашеная капуста. Далее промываем емкость и, когда она слегка обсохнет, насыпаем в нее 2-3 кило негашеной извести (в зависимости от объема, для маленького бочонка достаточно 1 килограмма).
Наливаем горячую воду, постепенно доведя ее уровень почти до краев, и накрываем крышкой, после чего выжидаем около 3 часов. Затем опорожняем емкость и несколько раз споласкиваем чистой водой. Для мытья больших бочек лучше всего использовать круглую щетку на длинной рукояти, особенно в тех случаях, когда нужно промывать емкости известковой водой.
Хранить деревянные дубовые бочки и кадки нужно обязательно наполненными водой, чтобы они не рассыхались. Тем более, что у современной продукции бондарей обручи, как правило, сделаны из нержавеющей стали, и сырость им не страшна. Зато древесина, пропитанная водой, набухает и хорошо удерживает жидкость. Что примечательно, чистая питьевая вода в дубовых бочках и жбанах никогда не портится. Впрочем, если вы вдруг обнаружили забытую пустую бочку и нашли у нее небольшие зазоры между досками, не торопитесь избавляться от емкости. Достаточно снова залить воды на сутки, и древесина снова разбухнет, сомкнув щели.
Подготовка бочонка из дерева
Подготавливая деревянные бочки для закладывания в них солений, повторите операцию с негашеной известью, если вы ощущаете затхлый запах. Но прежде чем выливать получившееся известковое «молоко» по истечении 3 часов, нужно этим раствором вымыть дно и стенки. Только потом прополощите несколько раз емкость, все время меняя воду на чистую. Если вдруг обнаружена плесень, лучше засыпать в ведро воды 100-200 граммов каустической соды или золы, получив в итоге щелок, которым и отмывается бочка или жбан, после чего тщательно пропаривается и споласкивается чистой водой.
Если банки перед закруткой солений или варений стерилизуются на пару или в микроволновой печи, то с бочкой такую операцию выполнить невозможно. Зато дезинфекцию можно выполнить иным способом. Для этого берем хлорную известь и разводим ее в воде, из расчета 100-200 граммов на каждые 10 литров. Затем полученным раствором промываем бочку. В результате вы добьетесь идеальной чистоты, но емкость будет пахнуть очень неприятно. Чтобы избавиться от запаха, несколько раз заполняем бочку горячей водой, которая сливается, когда остынет.
Совсем иначе надо поступить с только что купленной бочкой. Прежде всего, позаботьтесь о приобретении как минимум за месяц до начала консерваций. Новую емкость залейте водой до верха и держите наполненной не менее 2 недель, опорожняя и снова заливая до краев каждые 5 дней. Далее подготовьте большое количество кипятка. На дно бочки уложите свежесрезанные ветки полыни и можжевельника. Залейте емкость закипевшей водой и накройте крышкой, после чего укройте одеялом. Чтобы вода долго не остывала, калим на огне большие булыжники и кидаем периодически в бочку, чуть приподняв крышку. Опустошив, выставляем на свежий воздух, а перед как засолить помидоры или огурцы, нужно сжечь на дне немного серы, опять же накрыв крышкой.
Засолка огурцов в дубовой емкости
После сбора урожая нужно тщательно перебрать огурцы и выбрать те, которые еще не совсем созрели, поскольку в деревянной бочке они быстро дойдут до нужной степени зрелости. При этом откладываем для засолки только одинаковые по размеру. Это одно из основных правил при консервации в бочке, чтобы овощи были отсортированы по величине, что необязательно при закрутке в стеклянную тару. Замачиваем огурцы в холодной воде не менее чем на 2,5 часа (бочку для этой цели не используем), если же к засолке готовятся купленные овощи, лучше удвоить время.
Рекомендуется использовать емкость из дуба или липы объемом около 10-20 литров. Дело в том, что чем больше кадка, тем хуже получается соление.
Засолка огурцов в бочке
Далее, перед тем, как солить огурцы, стенки и дно в бочке натираем очищенным и мелко порубленным чесноком (100 граммов), который предварительно перетирается с солью. Затем на дно, полностью его закрывая, бросаем листья хрена, 100 граммов листьев смородины и столько же нарезанного соломкой хрена. Туда же помещаем 0,5 кило порубленного 10-сантиметровыми отрезками укропа и 50 граммов нарезки эстрагона, а также 10 граммов нарезанного кольцами горького перца. Затем плотно, слоями, устанавливаем вертикально огурцы, по мере заполнения бочки помещаем между овощами и стенками листья хрена по всей окружности.
В середине укладки помещаем столько же специй и пряностей, сколько в начале, и оставшуюся треть – сверху, снова накрыв хреном. Для рассола берем для мелких и средних огурцов около 70 граммов соли на литр воды, если же овощи крупные, нужно взять соли на 10 граммов больше. На каждые 10 кило огурцов вам понадобится приблизительно 2 литра заливки, которую перед заполнением бочки процеживают через марлю. Емкость накрываем крышкой или кружком с гнетом. Хранить соленые огурцы лучше в прохладном месте, консервирование же происходит при комнатной температуре.
Солим помидоры в деревянной кадке
Как и в предыдущем рецепте, все начинается с сортировки, поскольку овощи должны быть одинаковыми по размеру. Разносортица также не нужна. Перед тем, как солить помидоры в бочке объемом 10 литров, кладем листья хрена и смородины так, чтобы они полностью покрыли дно. Сверху высыпаем нарубленный соломкой корень хрена вперемешку с крупной нарезкой укропа. Чеснок не используем, поскольку он может послужить причиной закисания помидоров из-за наличия в составе сахара. Начинаем закладывать чисто вымытые помидоры плотно, можно на бок, избегая деформации и разрывов кожицы.
Соление помидор в деревянной кадке
Перед укладкой второго ряда слегка перетряхиваем из стороны в сторону бочонок, чтобы овощи, уже помещенные в него, легли плотнее. Когда емкость заполнится почти доверху, насыпаем сверху новую порцию нарезки укропа и корня хрена, по объему такую же, как и первоначальная. Накрываем все снова листьями хрена и смородины. Для заливки в холодной воде путем тщательного перемешивания растворяем 60 граммов соли на каждый литр. Собственно, на 10 литр консервации именно столько раствора и понадобится. Соление помидоров в бочке происходит под гнетом и с закрытой крышкой в течение полутора месяцев.
Оцените статью: Поделитесь с друзьями!distiller
Из всех бондарных изделий была, есть и остается наиболее распространенной бочка, которая чаще всего бывает с выпуклым остовом. Для создания деревянной бочки используют дощечки-клепки, или лады. Из них, в свою очередь, формируют три набора. Для изготовления первого главного набора, предназначенного для боковой стенки, или остова бочки, используют изогнутые длинные и узкие дощечки-клепки. Два других набора представляют собой днища, или донья, плоской формы, большей частью округлые. Для того чтобы днища держались в ладах, в последних в обоих концах выбирают фальц, называемый уторным пазом, или просто утором. В него входят поперечные доски, составляющие донья. Сами боковые дощечки (клепки, лады) отфуговывают по боковым граням так ровно, чтобы они прилегали друг к другу очень плотно. Этому плотному прилеганию помогают стягивающие их обручи — железные или деревянные.
Бочка, по В.И.Далю (от «бочковатый», «бочистый», «бок»), — вязаная обручная деревянная посудина, состоящая из ладов, или клепок, двух дон, врезанных в уторы, и обручей. Понятно, что свое название эта деревянная посудина получила из-за выпирающих в стороны боков. Кстати, эта конструктивная особенность бочки с выпуклым остовом (в отличие от прямого) придает ей особенную прочность. В больших бочках, если необходимо, просверливают дыру, в последнюю вставляют кран (верток) или затыкают так называемым гвоздем (затычкой).
Открытые бочарные изделия (кадушки, ведра, ушаты, чаны и т.д.) имеют одно дно. Их боковые остовы представляют собой прямые стенки, располагаемые под острым, прямым или тупым углом по отношению к плоскости дна.
Размеры длины клепок и днищ у бочек колеблются от 60 до 180 см. Для клепок длиной 180 см берут кряж соответствующей длины (с прибавкой 4—5 см), диаметром 40— 50 см. Из такого кряжа должно выйти 24 клепки шириной 14—16 см и толщиной 4 см.
Для клепок длиной 150 см берут кряж, имеющий в диаметре 36—40 см. Число клепок из такого кряжа получают 24, ширина каждой 10 см, толщина 4 см.
Для клепок длиной 120 см и 90 см подойдет кряж диаметром 28—36 см. Ширина клепок 8 см, толщина 3 см.
Для клепок длиной 60 см кряж берут диаметром 18—26 см. Ширина полученных клепок будет 6—8 см, а толщина 1,5—2 см.
Кряж размечают. Затем каждую шестую часть делят на четыре. Из них уже делают клепки нужных размеров, следя за тем, чтобы заболонь и сердцевина были отколоты. В том случае, когда кряж будет больше нам необходимого для выделки соответствующего размера клепок, его можно разметить и другим способом — двухрядным или трехрядным.
Для распиловки кряжа на клепки можно предложить следующие схемы.
Для днищ 180-сантиметровых бочек идет кряж диаметром 56—60 см, длиной 94 см. Ширина досок 30 см, толщина 3—4 см.
Чтобы изготовить 40-ведерную дубовую бочку, нужны клепки длиной 90—120 см, шириной 8—14 см, толщиной 2—3 см.
Для обычных кадок заготавливают клепки 60—90 см длиной, 8—12 см шириной. 4 см толщиной.
Для небольших бочонков и ведер изготавливают клепки длиной 60—90 см, шириной 10 см и толщиной 2—3 см.
Наиболее ходовыми бочками являются такие, у которых высота составляет 50 и 70 см. В целях более экономного расхода материалов есть смысл изготавливать бочки по парам. Одну высотой 50 см, другую высотой 70 см. В этом случае отходы более крупной бочки могут послужить заготовками для мелкой.
Из-за яйцевидной формы расчет объема бочки затруднителен. Однако на практике бондари нашли способ быстрого и довольно точного подсчета этого объема. Итак, для расчета объема бочки необходимо измерить ее высоту от одного утора до другого, а также диаметры в двух местах: в центральной части и в дне. Измерения лучше производить в дециметрах (напомним, 1 дм= = 10 см), так как 1 дм3 равен 1 л. Затем каждый измеренный диаметр возводят в квадрат.
Далее большее из полученных чисел удваивают и прибавляют к меньшему. Результат умножают на высоту бочки, а потом еще раз умножают на 3,14. Полученное от умножения произведение делят на 12 и получают объем бочки в литрах. Чтобы узнать, какое количество ведер содержится в бочке, ее объем в литрах делят на 12 (обычный объем’одного ведра в литрах).
Для примера рассчитаем объем бочки, у которой высота 70 см (7 дм), большой диаметр 60 см (6 дм), малый диаметр (поперечник днища) 50 см (5 дм). Произведем расчеты:
1) 5×5 = 25 дм2;
2) 6×6 = 36 дм2;
3) 36 х2 = 72 дм2;
4) 72 + 25 = 97 дм2;
5) 97 дм2 х7 дм = 679 дм3;
6) 679 дм3хЗ,14= = 2132 дм3;
7) 2132 дм3 : 12 = 148 дм3 = = 148 л;
8) 148 л : 12 = 15 ведер.
В буквенном выражении формула расчета объема бочки будет выглядеть так:
(d2 + 2D2) • h — пгде: V — емкость бочки в литрах;
d — диаметр дна бочки;
D — диаметр центральной части бочки;
h — высота бочки;
л — постоянная величина 3,14.
Какой формы и сколько нужно клепок?
Для облегчения поиска ответов на поставленные вопросы бондарь на листе картона или бумаги чертит окружности центра и дна будущей бочки. Причем можно чертить в масштабе 1:1. Тогда расчеты упрощаются. Или можно чертить с соответствующим уменьшением в 2, 4, 5 раз и т.д. И тогда при расчетах необходимо это уменьшение учитывать.
Итак, мы знаем, что в нашем примере большой диаметр равен 60 см. Диаметр дна 50 см. Вычерчиваем соответствующие диаметры на чертеже. Если нам известен только диаметр дна, то без особого труда (прибавив 1/5 часть диаметра дна) можно получить диаметр центральной части бочки (брюшной). И наоборот. Если нам известен большой диаметр, то можно вычислить (отняв 1/6 большого диаметра) диаметр дна.
Установить число клепок можно двумя способами. Либо, зная ширину в центре одной заданной клепки, выстраиваем на чертеже по большой окружности необходимое количество данной величины. Либо данную окружность делим на определенное число раз (в нашем случае на 16) и таким образом узнаем ширину самой широкой части клепки. Зная радиус большой окружности (30 см), по известной формуле (2tcr) находим длину этой окружности: 2x30x3,14 = 188,4 см.
Теперь эту длину делим на количество клепок (16). Получим 11,7 см. Округлив это число, получим 12 см. Такова будет ширина центральной части клепки. Если на чертеже мы проведем соответствующее число радиарных линий (в нашем случае 16), то здесь же на чертеже сможем измерить ширину конца клепки. Она составит приблизительно 10 см. То есть ширина конца клепки будет меньше ширины центральной ее части на 1/6 последнего размера.
Мы можем установить также и кривизну (выпуклость) клепок и величину скоса боковых граней. Мы можем количество клепок увеличить или уменьшить. Соответственно изменятся и размеры каждой отдельно взятой клепки. Заметим, что при заданной высоте бочки 70 см от утора до утора действительная длина клепки должна быть примерно 84 см (с учетом на изгиб и на обрезку).
Толщина клепки во взятом примере будет равна 2 см (60—50 = 10 см; 10:5 = 2 см). Толще V — обший объем цилиндрического изделия; d — диаметр дна; h — постоянная величина, равная 3,14.
Внутренний объем конических бондарных изделий вычисляют по формуле усеченного конуса:
V = л h (D2 + d2 + Dd).
Буквенные обозначения в данной формуле прежние.
Выделка клепок, или ладов
Расскажем о выделке клепок пооперационно.
1. Нарезка клепок. Для выделки клепок используют разные породы деревьев. В зависимости от предназначения бочек выбирают и соответствующее дерево. Скажем, лучшими бочками считаются дубовые. Их в основном предназначают для хранения спирта, коньяка, пива, вина и т.п. Для изготовления клепок под бочки, используемые в виноделии, идет обычно белый дуб.
Кстати сказать, использование дубовых бочек в виноделии очень часто является необходимым технологическим условием для получения соответствующего напитка. Так, например, спиртной напиток ром (крепостью 45%) получают из выдержанного ромового спирта, который происходит в результате сбраживания и перегонки сока сахарного тростника. Выдержка рома в дубовых бочках является непременным условием технологии.
Если в бочке собираются хранить воду, то для нее клепки делают из сосны, осины или ели. Для хранения молока и молочных продуктов на бочки идет можжевельник и липа.
К исходной древесине предъявляют определенные требования. Она должна быть сухой и без пороков: без отлупа, червоточины, прорости, свилеватости, исззаросших сучков, без так называемых раковин. О гнилом и прелом дереве и говорить нечего. Ясно, что таковое для выделки бочек не годится.
Для изготовления клепок лучше всего брать лес, колотый по сердцевинным слоям. Клепки из таких дощечек получаются наиболее прочными на изгиб. Обычно их вытесывают специальным бондарным топором. Но делают клепки и пилеными. Если вытесненные клепки предназначают для бочек, в которых потом собираются хранить различные жидкости, то пиленые клепки идут на бочки для сыпучих материалов — песок, мука и т.п.
Колоть клепки лучше всего из дерева, только что срубленного. А время заготовки наиболее подходящее — это октябрь и ноябрь. Деревья при помощи пилы или топора валят на землю. И тут же разделывают на клепки. То есть вначале дерево очищают от сучьев, затем распиливают на кряжи с таким расчетом, чтобы они по Алине на 2—3 см превышали будущие клепки или даже еще больше. Далее кряжи колят по сердцевинным лучам на части. Иногда колят и по годичным кольцам. Тогда клепка получается уже выпукло-вогнутой. Но по сердцевинным лучам колоть легче. Удобно колоть топором-колуном, у которого обух толстый, а клин острый и широкий.
В зависимости от толщины кряж колят вначале на половины, затем на четвертины, на восьмые доли. Если возможно, то колят и на шестнадцатые доли и т.д. От полученной минимальной части кряжа откалывают заболонь и сердцевину — то есть самые рыхлые слои древесины вместе с корой при помощи клиновидного изогнутого ножа. Теперь полученную среднюю часть колят по годичным кольцам надвое или натрое. Вновь полученные части называют гнатинником. По ширине его стараются получить на 1 см больше ширины будущей клепки. А вот уже гнатинник разделывают на клепки. Ясно, что и по толщине заготовки должны превышать толшину будущей клепки: ведь сырой лес, высыхая, на 12— 20% сократится. Бондарь по опыту знает, каких размеров ему делать заготовки в зависимости от породы и влажности леса.
Мы уже видели схемы однорядной, двухрядной и трехрядной выколки кряжей. Заметим, что больше всего отходов получается при однорядной выколке.
Сухое дерево колется труднее. Пилить клепки, естественно, из сухого дерева легче. Клепки нарезают с таким расчетом, чтобы в середине они были шире, чем по концам (точнее, их потом стесывают). Но по концам их толщина несколько больше, чем в средней части. Утолщение по концам необходимо для вырезки потом утора, то есть паза под дно или днище. Для правильной и более быстрой нарезки клепок используют шаблон. В качестве последнего может послужить уже готовая клепка. Можно также изготовить шаблон из фанеры в виде готовой клепки.
2. Сушка клепок. Прежде чем окончательно отделывать клепки, их сушат. Клепки складывают по две крест-накрест. Естественная сушка может длиться до года. Поэтому обычно бондарь делает себе запас клепок на это время. Сушить клепки можно и в специальной сушилке — закрытом помещении с подогревом и с обеспечением циркуляции воздуха.
Если бондарь делает бочки, как говорится, для своей надобности, то необходимость в устройстве специальной сушильни отпадает. Ведь для изготовления одной-двух бочек клепки можно высушить и дома над печкой или без нее, если дом не сельский и не дачный. При сушке следят за тем, чтобы клепки не растрескались, особенно на торцах. Для этого последние смазывают глиной или краской или еще заклеивают бумагой. По времени сушка может длиться от одного дня (например, на жарко истопленной печи) до нескольких дней (в теплом помещении).
3. Обработка клепок.
После сушки доски как клепок, так и днищ обрабатывают, то есть им придают именно ту форму, которая необходима для изготовления бочек.
Обычно клепки делают на 2—3 см длиннее, чем необходимо, поэтому после сушки их укорачивают с обоих концов лучковой пилой. Если бочку делают с вогнутым дном, то клепки не укорачивают, а спиливают, выравнивают в седле, когда бочка собрана, связана обручами и уже намечено место для дна.
Просушенные и укороченные клепки обрабатывают внутри и снаружи. Обрабатывает их каждый бондарь по-своему. В результате обработки клепки должны быть очень точно подогнаны друг к другу.
Вначале обработки клепку стесывают с наружной стороны специальным бондарным топором (он сточен на одну сторону). Работает бондарь на чурбане, держа клепку левой рукой и производя стесывание правой. Можно стесывать не только топором, но также одним из стругов или косарей на бондарной скамье. Движения бондаря при этой работе должны быть неторопливыми, очень расчетливыми, с тем чтобы не испортить клепку излишним отщепом или надрубом. Как правило, косарями, шляхтами и стругами бондарь пользуется для последующей чистовой обработки клепки. Обтесанную снаружи и внутри клепку сверяют с шаблоном. Когда обтесывание закончено приступают к обстругиванию клепки. Для этой цели сначала берут рубанок с выпуклой подошвой и с лезвием дугообразной формы Им обстругивают клепки, а затем слегка сглаживают последние прямым рубанком, снимая небольшую стружку. Окончательную отделку и обработку клепок осуществляют тогда, когда они уже собраны в бочку. На ,с показана клепка такой формы, какой необходима для изготовления выпуклых бочек. Форма может быть, Эта клепка в середине гораздо шире, чем по краям. Скашивают клепку к краям очень аккуратно. Работу эту можно делать на глаз, но лучше, все время сверяя с шаблоном, отмечая неровности карандашом. В выполнении данной работы нужна не только аккуратность, но еще и большая точность. Если ее не будет, то при сборке бока клепок могут не сойтись, и хлопот тогда по пригонке не оберешься.
О внутренней обработке клепки скажем несколько подробнее. При этой работе прежде всего намечают толщину клепки по всей поверхности, особенно старательно в шейках, то есть на концах. Толщину отмечают при помощи шаблона — чертилки. Чертилку накладывают в середине клепки так, чтобы острие а пришлось на самое ребро клепки. Затем шаблон ведут по всей длине клепки. Острие б отметит толщину шейки. Понятно, что при изготовлении бочек разных размеров разными будут и толщины клепок. А следовательно, и чертилок у бондаря должно быть несколько. Клепку с размеченной толщиной укрепляют в станке и всю лишнюю древесину стесывают топором или стругом.
Последняя операция по обработке клепок — это их прифуговка. Как мы уже говорили, очертания будущей бочки напрямую связаны с формой клепки. Если у клепки боковые линии будут прямыми, то и бочка получится прямой. Самая прочная и удобная форма бочки выпуклая. Для нее клепку делают такой. То есть у нее середина широкая, концы сужены. Наиболее распространенное соотношение середины и концов клепки, как мы уже отмечали, следующее: на конце клепка должна быть уже или меньше середины на 1/6 часть. К примеру, если в середине ширина клепки 12 см, то в концах она будет равна 10 см. Соотношение может быть и другим. Заметим, чем больше разница между шириной в середине и в конце клепки, тем круче будет бочка в боках.
Обстругивают и прифуговывают размеченные ребра клепки рубанком и фуганком, закрепляя ее в ладиле. А можно эту операцию выполнять и на большом бочарном рубанке. При фуговании ребра не прифуговывают вплотную, а делают небольшой зазор. То есть ребра клепок чуть скашивают внутрь. При стягивании бочки обручами имеющийся зазор исчезнет: клепки плотно прижмутся друг к другу.
Днища
Эти части бочки изготавливают из досок, которые чуть толще клепок. Доски обстругивают сначала рубанком, а затем прифуговывают плотно друг к другу. В зависимости от ширины досок и размера бочки днище может быть сколочено из четырех, пяти, шести и т.д. досок. Удобнее дощечки для днища нарезать из одной доски. Так как днише у бочки имеет круглую форму, то и составные дощечки подбирают такой длины, чтобы потом при придании днищу округлости было меньше отходов. Строгают доски днища, как правило, снаружи. Изнутри либо совсем не строгают, либо строгают только слегка.
Обручи
Их делают либо железными, либо деревянными. Железные изготавливают из полосного железа, ширина которого зависит от величины бочки. Чаще всего ширина составляет 3—4 см. Концы полосного железа накладывают друг на друга и склепывают. Железные обручи целесообразно применять для крупных бочек. Для деревянных обручей используют древесину клена, дуба, вяза, бука, ясеня. Применяют для деревянных обручей и какое-либо другое прочное и гибкое дерево — можжевельник, черемуху, ель и т.д. Для обручей выбирают молодое дерево, которое подрезают каждые 10—12 лет, — оно наиболее гибкое. При заготовке древесины для обручей пользуются следующими инструментами: топор, нож, струг, мялка, щепальные клинья, или колонки. Хорошо заготавливать деревянные обручи поздней осенью или в начале зимы. Кору с молодых деревцев или прутьев не снимают. В зависимости от толщины каждый прут расщепляют вдоль на две половины, на три или на четыре части.
Чтобы расщепить на две пластины, удобно пользоваться ножом. В остальных случаях пользуются щепальным клином, изготовленным из твердых пород дерева. В пруту делают надрез ножом на три или четыре части. Вставляют в надрез соответствующий щепальный клин и натягивают на него прут. Последний расщепляется на нужное нам количество частей. Чаще всего обручи делают из половинок прута, которые сгибают вокруг вбитых в землю по кольцу кольев. Концы обручей заводят за колья. Зафиксировав таким образом обручи, им дают высохнуть. Но удобнее для гнутья обручей пользоваться специальной конусообразной болванкой. Верхняя часть этой болванки соответствует малым обручам, нижняя — большим. Иногда заготовки, перед тем как согнуть в обручи, распаривают. Для удобства гнутья пользуются вспомогательными инструментами — мялкой или специальной скобой вбитой в стену либо в деревянный брус.
Сборка клепок
После того как подготовлены клепки, днища, обручи, приступают к сборке бочки. В первую очередь, конечно, собирают клепки. Но, перед тем как собирать их, клепки необходимо, по выражению бондарей, причертить друг к другу, то есть подогнать, прижать. Причерчивают при помощи обычного циркуля, рейсмуса или штангенциркуля. Отыскивают на концах каждой клепки середину и отмечают ее. Далее находят середину по длине клепки и, поставив здесь острие неподвижной ножки циркуля, другим концом очерчивают дугу на концах клепки. Проделав эту операцию со всеми клепками, находят таким образом шейную линию. Именно по ней потом придутся уторы для вставки днищ.
После расчерчивания приступают к сборке клепок. Берут вначале головной, или торцевой обруч (тот, которым стягивают клепки по концам) и прикрепляют к нему втулочную клепку. Таковой называют ту клепку, в которой будет находиться втулка бочки, если она запланирована. Втулочную или обычную первую клепку прикрепляют к обручу при помощи струбцинки или зажима, похожего на бельевую прищепку.
Оговоримся, в бондарных мастерских начинают сбор остова бочки при помощи специального рабочего обруча. Он представляет собой металлическое кольцо из круглого или полосового железа толщиной 10— 15 мм. Диаметр рабочего обруча обычно несколько больше диаметра постоянного — ведь его потом убирают, заменяя последним. В зависимости от размеров бочки в бондарных мастерских имеется несколько рабочих обручей, которые дублируют постоянные (головные, они же шейные или торцевые, срединные, или брюшные). Используют также и предохранительный обруч, который, в сущности, тот же рабочий.
Итак, продолжим речь о сборке клепок в остов. Прямо напротив первой клепки ставят самую широкую или основную клепку, а между ними по бокам на одинаковом расстоянии еще по одной. Клепки также закрепляют струбцинками или зажимами. Подобная расстановка клепок поможет прочно удержать головной обруч как бы на четырех ножках. Далее остальные клепки расставляют на свои места. Затем снимают зажимы и головной обруч несколько осаживают книзу, одновременно нагоняя на остов один или два шейных обруча и один средний (его еще называют брюшным, или пуковым). Можно эту первоначальную работу по сбору клепок в остов проделать иначе. То есть, поставив две клепки напротив друг друга, накладывают обруч и устанавливают другие клепки одну за одной, прикрепляя зажимами. Конечно, трудно приготовить клепки, которые подошли бы друг к другу, как говорится, без сучка и задоринки.
Случается так, что последняя клепка оказывается более широкой, чем необходимо. Тогда одну или две соседние клепки уменьшают в ширине. Или одну широкую заменяют двумя узкими клепками. В том случае, если не совпадают диаметры краев бочки, то есть один край шире или уже другого, две-три или несколько клепок перемещают концами в обратную сторону. Таким образом добиваются равенства диаметров у верхнего и нижнего основания бочки. Когда все клепки расставлены, надеты шейный и срединный обручи, остов переворачивают и стягивают клепки при помощи ворота или веревки. Однако стягивать клепки нужно осторожно, чтобы не поломать какие-либо из них. Лучше всего стягивать предварительно распаренные клепки. Есть несколько способов нагрева и распаривания последних. В больших бочарных мастерских пользуются специально устроенной мангальной печью с жаровым колпаком. Для мастерских меньшего размера можно порекомендовать железный таганчик-мангалку. Производят распарку клепок при помощи еще железной круглой печи с надставной трубой.
Дупло (так бондари называют полусобранный остов) надевают на эту печь. Ее топят, а клепки с внутренней стороны предварительно смачивают водой. При нагревании происходит распаривание клепок. После этого они становятся более податливыми на изгиб, менее ломкими. Если диаметр бочки меньше нашей круглой печки, то дупло надевают на печную трубу, предварительно сняв у нее одно колено, а потом (после размещения дупла), поставив его на место. Теперь печная труба, проходящая через дупло бочки, будет производить нужную нам по распариванию работу. Само дупло размещают на подставах, сверху и снизу прикрывая железными крышками. Каждую из крышек вырезают из листового железа в виде двух полукругов с аналогичными полукруглыми вырезами для пропуска печной трубы. Опять обильно обрызгивают дупло водою перед распаркой, да и во время оной. Вода от жара печной трубы нагревается, превращаясь в пар. Ну а последний делает свое распаривающее дело. Сколько распаривать клепки — каждый бондарь решает опытным путем. Обычно эта операция длится 1—2 часа. Слишком распаренные клепки становятся чересчур мягки на изгиб. Недораспаренные клепки при сгибе лопаются.
Продолжительность распаривания зависит также и оттого, насколько клепки необходимо согнуть. Если мы изготавливаем небольшую бочку с малым изгибом клепок, то необязательно прибегать к помощи железной круглой печи. Можно воспользоваться и железным таган-чиком-мангалкой. В мангалке разжигают дрова. Когда образуются жарко тлеющие угли, ее ставят в середину дупла и производят распаривание клепок. Разумеется, эту работу проделывают в каком-либо нежилом помещении, где существует свободный обмен с наружным воздухом. Распаренные клепки стягивают. Делают это, как уже отмечали, при помощи затяжек и ворота или при помощи обычной палки и веревки (закрутка). На шейную часть остова набрасывают веревочную петлю и постепенно ее затягивают. Если имеющиеся клепки толстые (как правило, у больших бочек), то используют не одну, а две, а то и три затяжки. Затягивают постепенно. Сначала стягивают срединную часть, затем шейную. Полезно дупло бочки крутонуть то в одну, то в другую сторону, поворачивая как рулевое колесо у автомобиля. Это помогает стяжку клепок сделать равномерной. Порою то одна, то другая клепка выпирает из общего ряда. Ее вправляют при помощи деревянного молотка — киянки. Когда концы клепок достаточно плотно сойдутся, на дупло бочки нагонять начинают обручи. Вначале большой (брюшной), затем шейный и головной. Эти обручи рассматриваются как рабочие. Постоянные обручи нагоняют на бочку после вставки днищ.
После того как клепки стянуты с одной стороны дупла, его переворачивают и дотягивают клепки на другом конце. Полученный предмет со стянутыми клепками уже по праву называется остовом бочки, или бездонной бочкой. Этот остов с рабочими обручами сушат в течение нескольких дней или одной-двух недель (в зависимости от условий сушки: около печки или на открытом воздухе). Затем его закаливают изнутри, то есть подвергают обжигу. Для этого в остове разжигают стружки. Далее остов перекатывают, следя за тем, чтобы древесина не обуглилась, а лишь слегка подкалилась, приобретя золотистый оттенок. Так поступали старые мастера. Но проще подвергнуть остов опаливанию паяльной лампой, соблюдая, естественно, правила противопожарной безопасности. Обжиг или закаливание производят дня того, чтобы клепки в остове стали значительно устойчивыми по форме. В промышленных условиях закаливание производят на мангааьной печи. Небольшие бочонки можно и не обжигать. Их достаточно высушить при высокой температуре, например, в духовке русской печи.
Остовы конической формы (с прямыми стенками) вообще не закаливают, так как их клепки не имеют изгиба по длине. После закалки бездонной бочки ее обручи осаживают, так как во время обжига древесина размягчилась, часть ее влаги испарилась, то есть клепки несколько усохли. Осаживают обручи при помощи молотка и набойки. Во время этой операции клепки своими ребрами плотно прижимаются друг к другу, не оставляя никаких щелей или зазоров. Все неровности просто-напросто сминаются. Затем приступают к обрезке выступающих концов клепок лучковой пилой, расположив остов в седле или на скамье.
Заметим только, что обрезку ведут так, чтобы поверхность пропила была на клонена несколько внутрь остова. Далее снимают фаски при помощи бондарного ножа, струга или бочарного рубанка. Съему фасок или срезов производят на половину толщины торцов. Таким образом, предупреждают какие-либо откалывания концов клепок, их расщепливание на внутренней стороне остова. Торцы последнего после съемки фасок вообще приобретают опрятный и красивый вид. Здесь мы лишний раз убеждаемся в том, что красота и польза не разделимы, они очень тесно взаимосвязаны.
Снаружи края торцов мы пока не трогаем. Их отделку оставляем на потом, когда завершим изготовление бочки. Перед тем как вырезать уторы и вставлять днища, остов бочки остругивают изнутри и снаружи. Дело в том, что после обжига и осаживания обручей кромки соседних клепок часто образуют выступы (бондари называют их провесами). Вот эти провесы и необходимо сгладить при помощи стругов. Для наружного остругивания применяют вогнутый струг, скобель или рубанок, для внутреннего — выпуклый.
При остругивании снаружи временно снимают поочередно обручи. Сначала с одного конца остова, затем с другого. Особенно тщательно выравнивают шейную поверхность остова изнутри. Только в этом случае удается утор-ный паз выбрать ровным и по окружности и по глубине. А следовательно, и вставка днищ будет плотной и прочной. Иногда этой зачисткой шейной части на расстоянии 10— 15 см от края остова и ограничиваются.
После окончания зачистки приступают к выемке уторного паза. Эту операцию производят уторником. А если бочарное изделие небольшое и чистота, и правильность выемки не требуются, то уторный паз выбирают гребнем. И в том и в другом случае от края отступают 3—5 см.
Уторный паз выбирают только с одной стороны, если готовят бочку, открывающуюся с другого конца. Если предполагают изготовить бочку глухую, двухдонную (закрытую), то уторный паз выбирают в двух концах остова. Для совершения этой операции остов бочки размещают в седле или на верстаке. При выемке уторного паза бондари пользуются несложным правилом. Глубина паза не должна быть больше половины толщины концов клепок, а ширина утора не должна превышать толщину досок днища. Даже наоборот — ширину делают несколько уже толшины днища примерно на 3—5 мм. Так только можно будет добиться плотности прилегания днища в бочке и предотвратить возможную течь.
Теперь приступаем к изготовлению днищ. Хотя об этом речь уже шла выше, напомним, что днища делают из клепок-дощечек, разных по ширине, но одинаковых по толщине, плотно подогнанных и прифугованных друг к другу. Толщина днищ обычно превосходит толщину боковых клепок. В зависимости от величины бочарного изделия днища могут состоять из 4—6 дощечек, сплоченных в один щит. Прежде чем сплачивать дощечки в единый щит, каждую из них тщательно строгают стругом, скобелем, рубанком.
Также тщательно, а может, даже и более тщательно отфутовывают боковые грани. После этого дощечки зажимают в щемиле. Можно предварительно их сплотить при помощи шипов. На образовавшемся из дощечек щите, зажатом в щемиле, очерчивают круг будущего днища. Внимание — его диаметр должен превосходить диаметр бочки в уторе на двойную глубину уторного паза.
Теперь лишние части досок отпиливают лучковой пилой по сделанной разметке. Можно предварительно щит разобрать. А можно производить опиловку его прямо в щемиле. Внешнюю сторону днища еще раз тщательно остругивают. С внутренней стороны у днища стесывают края. Циркулем очерчивают границу этой отлогой фаски. Ее ширина обычно составляет 4—7 см.
Снимать эту фаску необходимо потому, что толщина досок днища больше толщины уторного паза. При снятой фаске днище зайдет в утор и по мере захода будет возрастать плотность соприкосновения его с уторным пазом. Иногда фаску снимают и с наружной стороны днища. Но фаску эту делают небольшой. По своей ширине она должна быть меньше глубины уторного паза. Тогда после вставки дна в бочку фаска полностью спрячется.
Доски, составляющие днище, каждая имеет свое название. В днище, состоящем из 4 досок, две срединные называются главными, а боковые — отрезами. В днище из 6 досок две средние также называются главными, следующие две — боковыми, а крайние — по-прежнему отрезами. Подготовленное днище вставляют в утор. Целиком днище вставлять трудно. Чаще его вставляют разобранными дощечками. Предварительно с конца остова бочки снимают один или два обруча.
Клепки при этом разойдутся. Вставляют днище, начиная с крайних (боковых) дощечек. Последнюю среднюю дощечку вставить труднее всего. Вставляют ее примерно в такой последовательности. Вначале заводят один конец в уторный паз. На другом краю отгибают одну или две клепки так, чтобы сподручно можно было завести и второй конец дощечки в утор. Совершая эту работу, пользуются вспомогательным инструментом: накидными клещами, натягом. Клепки при вставленном днище несколько разойдутся.
Их вправляют на место деревянным молотком. Вставив днище на одном конце бочки, вставляют аналогично и на другом. Второе дно вставить труднее, так как его уже нельзя поддержать снизу.
Не по одной дощечке, а целиком днище вставляют в следующем порядке. Сначала вводят в утор один торцевой край. Далее разводят широко клепки и вводят в утор днище целиком. Перед вставкой часто уторы промазывают с помощью лопаточки замазкой (смесь сурика или мела и вареного льняного масла — олифы). Для более плотного прилегания днища используют еще так называемую бочарную траву: ситник, тростник и т.п. Эту бочарную траву в уторном пазу размещают при помощи конопатки. После того как оба днища введены в уторы, клепки еще раз подправляют деревянным молотком, а потом их плотно стягивают с помощью затяжек. Завершают работу тем, что на концы бочки вновь надевают обручи.
Иногда для большей прочности днище бочки укрепляют наладной дощечкой — набойкой. Она представляет собой дощечку 15 см в ширину и 3—4 см в толщину. Длина ее соответствует диаметру днища. Набойку закрепляют поперек досок днища нагелями. Последние вколачивают в концы клепок рядом с уторным пазом. Нагели делают достаточно длинными с тем, чтобы крепление набойки было надежным. Форма нагелей необязательно должна быть круглой. Она может быть граненной, например четырехугольной. Даже лучше, если она будет таковой, так как при высыхании бочки круглые нагели порою выпадают, а граненные задерживаются. Число нагелей с каждой стороны набойки варьируется от 4 до 6.
Последняя завершающая операция по изготовлению бочек — набивка постоянных обручей. Число их различно. На большую бочку набивают до 18 деревянных обручей или 6—8 железных. Для бочки средних размеров обычное число деревянных обручей — 14—16 штук. Их градация следующая: 8 шейных (по 4 обруча с каждого края), 6 брюшных (по 3 обруча в половине бочки). Реже насаживают 10 деревянных обручей (6 шейных, 4 брюшных; и шейные и брюшные обручи поровну распределяются на обеих половинах бочки). Заметим сразу же, что бочка с 10 деревянными обручами менее крепкая, чем с 14.
Деревянные обручи изготавливают из обручных хлыстов. Этими хлыстами опоясывают бочку в том месте, где предполагают разместить обруч. Делают на хлысте и на бочке соответствующие пометки. На хлысте отмечают места зарубок для вязки замка. На замок оставляют припуск в 10—12 см с обоих концов обруча. Сами концы срезают наискось в виде заостренных языков. Там, где отмечены у нас были зарубки, делают надрезы на половину ширины обручного хлыста. На одном конце обруча надрез делают сверху, на другом — снизу. С внутренней стороны обруча, по направлению от надрезов к середине, делают выемки длиной 4—5 см, постепенно сходящие на нет. Теперь вяжут замок. А именно: концы обруча зацепляют друг за друга выступами надрезов, укладывают в соответствующие выемки. То есть концы заводят и прячут на внутренней стороне обруча. Часто обруч в месте вязки замка для прочности оплетают ивовыми прутьями.
С бочки, как уже понял читатель, рабочие обручи снимают, заменяя их постоянными. Делать это нужно последовательно: сначала заменяют брюшные обручи в одной половине бочки, затем шейные все в той же половине, а уж потом тоже самое совершают со второй половиной бочки. Последние шейные обручи особенно трудно натягивать на остов бочки. Обруч заводят на клепки сначала с одного края.
Затем с другого, помогая при этом себе натягом и затяжками. Натягом работают так. Конец его ручки упирают в бок бочки, на другой конец все той же ручки надавливают рукой. Обруч в это время захватом натяга слегка растягивается и, охватив концы клепок, стягивает их. Клепки постепенно одну за другой глубже заводят в обруч.
Иногда надетая полуокружность обруча соскальзывает с клепок. Чтобы этого не происходило, надетую половину обруча закрепляют на края остова небольшими гвоздями. Вбивать их следует не более чем на половину толщины концов клепок. После того как деревянный обруч натянут на бочку, его необходимо осадить на предназначаемое место.
Пользуются при этом деревянным молотком и набойкой. Набойку ставят углублением подошвы на ребро обруча. Ударами молотка по головке набойки обруч осаживают на место. Последний должен быть набит на бочку без каких-либо перекосов, до отказа, плотно охватывая’ ее окружность.
Изготовление железных обручей сходно с изготовлением деревянных. Ширина и толщина железных обручей зависят от размеров бочки. Обычно берут полосное железо 3—4 см шириной. Здесь также начинают работу с обмера бочки. Отрезают полосное железо с припусками с обоих концов обруча на накладку в 10—12 см. Ножницами или зубилом отсекают также уголки концов обруча. Затем эти концы либо сваривают, либо склепывают. Сварку можно производить так, как делали бондари в старину без сварочного аппарата.
В кузнице концы обруча раскаляли докрасна. А потом, не давая остыть, на наковальне, держа щипцами и ударяя кузнечным молотом, концы сваривали. Но чаще все же концы склепывают. Их накладывают друг на друга и высверливают либо пробивают минимум две дырки, отступая от края по длине обруча 2 и 6 см. Поверхности обруча при помощи молотка придают уклон с тем, чтобы он плотнее облегал окружность бочки.
Насаживают железные обручи аналогичным способом, как и деревянные. Только при этом пользуются уже железными и молотком и набойкой. Чтобы предупредить ржавчину, железные обручи окрашивают черной масляной краской. Вид у готового деревянного изделия с черными полосами обручей — загляденье.
После насадки постоянных обручей бочку отделывают окончательно. Проходят стругом или шлифчиком по днищам и бокам бочки. Обрезают концы бочарной травы около уторов, подчищают вылезшую из них замазку. Скобелем подправляют фаски. Если запланировано, то в бочке высверливают втулочное отверстие. Стенки отверстия делают либо отвесными, либо под наклоном.
Современные технологии бондарей ФранцииТаламически-вызванные синаптические взаимодействия в стволовой коре, выявленные с помощью оптических изображений
Abstract
Мы использовали оптическую визуализацию чувствительных к напряжению сигналов красителей для изучения пространственно-временного распространения активности в коре ствола мышей, вызванной стимуляцией таламокортикальных афферентов в подготовка срезов in vitro . Стимуляция таламуса при низкой силе тока приводит к тому, что активность в значительной степени ограничивается одним стволом и границей между слоями Vb и VI.Низкие концентрации бикукуллина, антагониста рецептора GABA A , увеличивают амплитуду оптических сигналов, не влияя на их пространственно-временное распространение. Более высокие концентрации бикукуллина приводят к пароксизмальной активности, которая распространяется по внутриколоночным и межколоночным возбуждающим путям. Повышение активности рецепторов NMDA путем удаления Mg 2+ из внеклеточного раствора резко изменяет пространственно-временную картину возбуждения: активность распространяется на супрагранулярный и инфрагранулярный слои и соседние цилиндрические колонки.Это усиленное размножение подавляется антагонистом рецептора NMDA AP5. Подобное усиление распространения активности может быть произведено путем стимуляции таламуса короткой высокочастотной серией импульсов. Применение AP5 подавляет частотно-зависимое распространение активности. Эти данные указывают на то, что пространственно-временное распространение активности в бочкообразной коре изменяется из-за изменения временных паттернов таламических входов посредством механизма, опосредованного рецепторами NMDA, и предполагают, что аналогичный процесс происходит во время повторяющейся активности взбивания.
Ключевые слова: соматосенсорная кора, чувствительные к напряжению красители, глутамат, NMDA, ГАМК, эпилепсия, временное кодирование, мышь
Обработка сенсорной информации в коре головного мозга может быть описана как иерархический процесс, в котором входы от «конкретных Ядра таламуса интегрированы своими постсинаптическими корковыми мишенями, которые, в свою очередь, обеспечивают входные данные для нейронов в других слоях того же функционального столба, а затем и в соседние столбцы (Gilbert, 1992; Keller, 1995; Rauschecker et al., 1997). Хотя эта иерархическая схема чрезмерно упрощена, поскольку в ней отсутствуют такие факторы, как параллельная обработка данных и взаимодействия с обратной связью, считается, что она достоверно представляет, по крайней мере, начальные этапы корковой сенсорной обработки (Stone et al., 1979; Felleman and Van Essen, 1991; Iwamura , 1993).
Чтобы понять механизмы, лежащие в основе корковой сенсорной обработки, необходимо определить, как таламические входы интегрируются их постсинаптическими кортикальными нейронами и распространяются по внутриколоночным и межколоночным кортикальным путям.Это требует методов для идентификации дискретных функциональных столбцов и выявления пространственно-временного распространения таламических входов внутри и между этими столбцами. В настоящем исследовании мы воспользовались функциональным подходом к визуализации (см. Орбах и Коэн, 1983; Чинелли и Зальцберг, 1987; Юсте и др., 1997; Келлер и др., 1998), чтобы изучить внутрикортикальную обработку таламических входов в соматосенсорной коре. и его модуляция глутаматергическими и ГАМКергическими процессами.
Соматосенсорная кора некоторых видов грызунов содержит представления мистициальных вибрисс (усов), которые взаимно однозначно отображаются топографически на соответствующие массивы дискретных клеточных агрегатов, называемых бочонками (Woolsey and Van der Loos, 1970).Нейроны в цилиндре слоя IV, который соответствует определенному усу, реагируют преимущественно на стимуляцию этого уса (для обзора см. Armstrong-James, 1995; Simons, 1995). Нейроны в супрагранулярном и инфрагранулярном слоях непосредственно над и под стволом образуют функциональный столбец клеток со сходными характеристиками ответа (см. Обзор Keller, 1995). Кроме того, отдельные бочки можно идентифицировать в неокрашенных срезах головного мозга in vitro из (Agmon and Connors, 1991; Gottlieb and Keller, 1997).В результате бочкообразная кора головного мозга является единственной областью коры, в которой можно идентифицировать функциональные столбы без использования электрофизиологических подходов, метаболических маркеров или функциональной визуализации. Эти преимущества делают ствол коры головного мозга отличной моделью для изучения иерархической обработки таламических входов с помощью внутриколоночных и межколоночных синаптических взаимодействий.
Некоторые из этих результатов ранее появлялись в абстрактной форме (Laaris and Keller, 1999).
МАТЕРИАЛЫ И МЕТОДЫ
Подготовка срезов. Протоколы животных, использованные в этом исследовании, соответствовали всем соответствующим институциональным и федеральным нормам. Молодых взрослых самцов мышей CD-1 в возрасте 21–30 дней анестезировали кетамином (30 мг / кг). Мозг удаляли и готовили таламокортикальные срезы толщиной 400 мкм, как описано Агмоном и Коннорсом (1991). Этот препарат сохраняет как вентробазальное (VB) ядро таламуса, так и соматосенсорную (бочкообразную) кору, включая функциональные связи между этими структурами. Срезы хранили в камере выдержки, которая содержала искусственную CSF (ACSF) при комнатной температуре, аэрированную 95% O 2 и 5% CO 2 .ACSF состоял из (в мм): NaCl 124, NaHCO 3 26, NaH 2 PO 4 1,2, KCl 3,2, MgSO 4 1,2, CaCl 2 2,4 и глюкозы 10.
Один час спустя отдельные срезы окрашивали чувствительным к напряжению красителем RH-155 (100 мкм; Molecular Probes, Eugene, OR). Краситель растворяли в ACSF, и один срез помещали в статическую баню, содержащую этот раствор, непрерывно обрабатывая 95% O 2 и 5% CO 2 в течение 30-60 минут.Затем окрашенный срез переносили в записывающую камеру погружного типа и непрерывно перфузировали ACSF со скоростью 2 мл / мин при комнатной температуре.
Электростимуляция и запись. Биполярный стимулирующий электрод, изолированный за исключением кончиков, помещали в таламус VB для активации таламокортикальных клеток (150–200 мкс). Импульсы постоянного тока подавались через оптически изолированный изолятор стимула (PSIU6; Grass, Quincy, MA), управляемый генератором импульсов (Grass S48).Генератор импульсов также использовался для запуска получения изображения аналого-цифровым (A / D) преобразователем (см. Ниже). Стеклянные пипетки (заполненные 2 м NaCl, 2–5 МОм) помещали в слой IV для регистрации потенциалов поля, вызванных таламом. Эти записи были оцифрованы в режиме онлайн и сохранены на компьютере Apple Macintosh.
Оптические записи. Методы, используемые для записи чувствительных к напряжению оптических сигналов, аналогичны методам, подробно описанным в других источниках (Salzberg et al., 1977; Wu and Cohen, 1993; Keller et al., 1998). Чтобы вымыть несвязанный краситель, окрашенные срезы перфузировали ACSF в течение не менее 15 мин перед началом оптической записи. Регистрирующую камеру устанавливали на стационарный вертикальный микроскоп (BX50WI; Olympus Optical, Токио, Япония), жестко закрепленный на виброизоляционном столе. Для питания вольфрамово-галогенной лампы мощностью 100 Вт использовался стабилизированный источник постоянного тока, и свет от этой лампы был ограничен по полосе с помощью интерференционных и тепловых фильтров; Если не указано иное, использовали полосовой интерференционный фильтр 720 ± 40 нм (Omega Optical, Brattleboro, VT).Свет, прошедший через препарат, собирали через водно-иммерсионный объектив 10 × (числовая апертура 0,3, Olympus) и проецировали на матрицу фотодиодов из 464 элементов (NeuroPlex; OptImaging, Fairfield, CT). Каждый фотодиод отбирал оптические сигналы из области ∼60 × 60 мкм 2 . Выходной ток каждого фотодиода отдельно преобразовывался в напряжение и усиливался в двух отдельных каскадах (1000 ×), мультиплексировался и оцифровывался с разрешением 12 бит с помощью аналого-цифрового преобразователя.Перед оцифровкой оптические сигналы фильтровались с частотой 500 Гц. Все электронные компоненты являются частью коммерческой системы NeuroPlex. Данные были собраны и сохранены на персональном компьютере, управляемом программным обеспечением NeuroPlex (OptImaging).
Чтобы точно идентифицировать области в срезе, из которых были собраны оптические записи, использовалось специально разработанное устройство светоделения (Microscope Services, Rockville, MD) для одновременного проецирования изображений среза и света от светодиодов. встроены в матрицу фотодиодов в плоскость изображения камеры CCD (CCD72; Dage, Michigan City, IN).Это позволило нам разграничить расположение отдельных бочек и ламинарных границ (рис.). Эти анатомические особенности, наблюдаемые на неокрашенных срезах, хорошо коррелировали с их появлением на срезах, окрашенных по Нисслю.
, Видеографическое изображение среза in vitro через кору ствола, полученное во время сеанса записи. Шестигранник разграничивает границу матрицы фотодиодов из 464 элементов. Кортикальные слои обозначены римскими цифрами .Обратите внимание на появление бочек в слое IV. Регистрирующий электрод ( e ) помещают в цилиндр слоя IV для регистрации потенциалов поля. B , Одновременная регистрация потенциалов поля и сигналов чувствительных к напряжению красителей в ответ на таламическую стимуляцию. В нормальном ACSF ( nACSF ) потенциал поля имеет две составляющие, а оптический отклик — единственную деполяризующую составляющую. Когда синаптическая передача подавляется в растворе, номинально не содержащем Ca 2+ (0 Ca 2+ ), второй компонент потенциала поля ( с ) и оптический ответ отменяются, что позволяет предположить, что оба они представляют постсинаптическую активность. корковых нейронов.Ранний компонент в потенциале поля ( a ) подавляется антагонистом каналов Na + TTX, что позволяет предположить, что он представляет собой потенциал действия пресинаптического таламокортикального соединения.
Если не указано иное, все записи были получены с частотой дискретизации 1,6 кГц. Изображенные оптические отклики представляют собой среднее значение пяти последовательных кривых, собранных с 20-секундными интервалами. Чтобы скорректировать пространственные различия в интенсивности освещения и длине пути света, сигнал, зарегистрированный от каждого детектора, был разделен на интенсивность света в состоянии покоя, рассчитанную для соответствующего детектора.Интенсивность света в состоянии покоя для каждого детектора рассчитывалась путем вычитания значений интенсивности, зарегистрированных при закрытом затворе, из значений, записанных при открытом затворе, когда не применялась стимуляция. Результирующие амплитуды сигналов выражаются как частичное изменение поглощения (Δ I / I ). Чтобы количественно оценить относительные изменения в поглощении света, мы рассчитали среднее значение и стандартное отклонение Δ I / I в течение 100 мсек, предшествующих стимулу; Амплитуды постстимульного сигнала выражаются как количество SD выше этих средних базовых значений.Оптические отклики, которые как минимум на 2 SD выше этого среднего значения, считались значимыми.
Анализ данных был проведен на компьютере Apple Macintosh с использованием процедур, разработанных в Игоре (WaveMetrics, Lake Oswego, OR). Для статистического анализа использовали непарный критерий Стьюдента t .
Фармакологические и ионные манипуляции. Фармакологические агенты готовили непосредственно перед использованием из исходных растворов и растворяли в ACSF. Следующие агенты были получены от Research Biochemicals-Sigma (Натик, Массачусетс): d (-) — 2-амино-5-фосфонопентановая кислота (AP5), бикукуллин метиодид (бикукуллин) и тетродотоксин (TTX).Номинально свободные от Ca 2+ растворы получали заменой Ca 2+ на эквимолярные концентрации Mg 2+ ; Номинально не содержащие Mg 2+ растворы получали путем исключения из растворов Mg 2+ .
РЕЗУЛЬТАТЫ
Происхождение оптических сигналов
Во всех экспериментах мы стимулировали таламус VB и регистрировали потенциалы поля и сигналы чувствительных к напряжению красителей от стволовой коры. Потенциалы поля, зарегистрированные в слое IV, показывают две составляющие: раннее начало (3-5 мс) низкоамплитудного отрицательного отклонения, за которым следуют одна или несколько более поздних и более крупных составляющих (рис.). Оптические сигналы имели один компонент с задержкой начала 5,27 ± 1,54 мс ( n = 84 среза) и относительно медленным временем нарастания (измеренным от начала до пика; 21,61 ± 5,56 мс; n = 84). Чтобы охарактеризовать происхождение записанных сигналов, мы фармакологически изолировали различные компоненты оптически записанных ответов и ответов потенциала поля. Чтобы подавить синаптическую передачу, мы промывали срезы в растворах, номинально не содержащих Ca 2+ . Это привело к полному подавлению оптических откликов и более поздних компонентов вызванных потенциалов поля ( n = 11; рис.). Напротив, ранняя составляющая потенциала поля не была затронута, но полностью исчезла после применения антагониста каналов Na + TTX (0,5 мкм; n = 4).
Эти находки показывают, что оптические ответы и поздние компоненты потенциалов поля полностью зависят от синаптической передачи. Это предполагает, что в наших условиях записи, оптических ответов представляют собой постсинаптические ответы в корковых нейронах.Пресинаптические, зависящие от Na + потенциалы действия таламокортикального соединения, очевидные в раннем компоненте записи потенциала поля, оптически не обнаруживаются. Это может быть связано с относительно низкой частотой дискретизации или объективом с низкой числовой апертурой, который мы использовали.
В дополнение к сигналам, связанным с красителями, оптические ответы могут также возникать из источников, присущих срезу (Grinvald et al., 1988; Yuste et al., 1997). Чтобы определить, вносят ли такие собственные сигналы вклад в формы волны, зарегистрированные в нашем исследовании, мы проверили зависимость этих сигналов от длины волны освещения.Когда освещение было изменено на длину волны вне спектра поглощения красителя (≥800 нм), оптический сигнал в коре головного мозга не обнаруживался. Это говорит о том, что оптические сигналы представляют собой реакции, связанные с красителем, а не являются результатом собственных оптических сигналов.
Оптические сигналы, связанные с красителем, также могут возникать в результате активации глиальных клеток. В этом случае ожидается, что оптические ответы, зарегистрированные от глиальных клеток, будут иметь медленный временной ход (> 1 секунды) по сравнению с ответами нейронов (Konnerth et al., 1987). Однако оптические сигналы, зарегистрированные в коре ствола, имели только один деполяризующий компонент, длительность которого составила 99,92 ± 21,34 мс ( n = 84; рис.). Эти данные предполагают, что связанные с красителем оптические сигналы, проанализированные в настоящем исследовании, отражают нейрональные реакции и не связаны с сигналами, исходящими от глиальных клеток.
Пространственное распределение оптических сигналов
Для анализа пространственного распространения оптических откликов, вызванных таламусом, мы создали карты с цветовой кодировкой, представляющие мгновенную амплитуду откликов, регистрируемых каждым фотодиодом (рис.). На этих картах амплитуда отклика представлена как количество стандартных отклонений над средними базовыми значениями, рассчитанными отдельно для каждого фотодиода (см. Материалы и методы).
Распространение оптически записанных ответов в коре головного мозга в ответ на таламическую стимуляцию. Каждая панель отображает амплитуду оптических откликов, записанных каждым из 464 фотодиодов. Амплитуды сигналов, выраженные как SD выше средних базовых сигналов, кодируются цветом и интерполируются. Временные интервалы (в миллисекундах), указанные слева внизу каждой панели, относятся к началу стимуляции.Панели в A, C, E и F отображают сигналы, записанные на пике откликов. На каждой панели нанесены ламинарные и бочкообразные полые границы, идентифицированные на видеографических изображениях и срезов, окрашенных по Нисслю. Пластинки обозначены римскими цифрами . A , Ответы на низкоинтенсивную (30 мкА) и высокую (100 мкА) стимуляцию таламуса VB. В обоих условиях отклики ограничиваются стволами слоя IV и границей слоев V – VI. Данные на этих панелях отображают максимальное пространственное распространение ответов. B , Распространение ответов на таламическую стимуляцию (20 мкА) в ACSF, номинально не содержащем Mg 2+ . Активность распространяется вертикально внутри цилиндрической колонны и горизонтально через супрагранулярный и инфрагранулярный слои. C , Ответы, зарегистрированные в том же срезе, изображенном в B, , в присутствии 1,2 мм Mg 2+ ( nACSF ) или в присутствии 0 Mg 2+ и антагониста рецептора NMDA AP5 ( + AP5 ). Распространение активности в условиях 0 Mg 2+ значительно ослабляется AP5. D , Распространение ответов на короткую серию таламических стимулов ( 4 × 100 Гц , 25 мкА). Обратите внимание на внутриколоночное и межколоночное распространение активности. E , Активность, зарегистрированная в том же срезе, что и в D , в ответ на однократную стимуляцию таламуса (25 мкА, 1 импульс ) или в ответ на серию стимулов в присутствии рецептора NMDA антагонист AP5 ( 4 × 100 Гц + AP5 ).Распространение активности в ответ на повторяющуюся стимуляцию значительно ослабляется AP5. F , Ответы на таламическую стимуляцию в nACSF или в присутствии бикукуллина антагониста рецептора GABA A (2 мкМ mBIC ). Относительно низкие концентрации бикукуллина увеличивают амплитуду, но не пространственное распространение ответов. G , Оптическая активность таламической стимуляции (при t = 0), зарегистрированная с одного фотодиода, расположенного в супрагранулярных слоях, в присутствии бикукуллина 5 мкм.За первоначальным постсинаптическим ответом ( стрелка ) следуют два последовательных пароксизмальных события ( звездочек, ). H , Распространение активности в ответ на таламическую стимуляцию (одиночный импульс, 20 мкА) в присутствии бикукуллина 5 мкм. За начальным откликом в слое IV и на границе слоя Vb – VI ( t = 20 мс) следует инициирование пароксизмального события в том же стволе ( t = 32 мс). Активность сначала распространяется вертикально на супрагранулярные слои, а затем на инфрагранулярные слои.Затем пароксизмальная волна распространяется горизонтально по отдельным полосам в супрагранулярном и инфрагранулярном слоях, в основном избегая слоя IV. Примерно через 100 мсек после того, как пароксизмальная волна отступит, вторая пароксизмальная волна появляется в инфрагранулярных слоях, проецируется на супрагранулярные слои и распространяется горизонтально по двум отдельным полосам.
Интенсивность стимула VB-таламуса была доведена до уровней, на 10% превышающих порог для обнаружения оптического ответа. Эти уровни стимула варьировались от 20 до 50 мкА и вызывали идентичный образец ответа во всех срезах ( n = 84).Чтобы идентифицировать локус этих ответов, мы наложили на карты активности рисунки ламинарных и бочкообразных границ, полученные из видеографических изображений и срезов, окрашенных по Нисслю (см. Материалы и методы; рис.). Это показало, что во всех случаях низкоинтенсивное таламическое моделирование вызывало реакции, которые были ограничены одной или двумя пустотами ствола и окружающими их стенками, а также областью аналогичного размера под этими стволами, рядом с границей между слоями Vb и VI ( Рис. А ).Пространственный разброс ответа был идентичным на протяжении всего периода ответа, и оптически зарегистрированная активность не распространялась за пределы областей, активированных изначально. Модели активности на Рисунке A отображают максимальное пространственное распространение ответов.
В большинстве (62%) срезов оптические отклики в слое IV и на границе слоя Vb – VI имели идентичные задержки начала. Реже ответы сначала появлялись на границе слоя Vb – VI (21%) или в слое IV (17%).В этих случаях разница в латентности начала составляла 4–9 мсек.
Увеличение интенсивности стимула расширило область, из которой были записаны оптические ответы, но эти области все еще были ограничены слоем IV и границей слоя Vb – VI (Рис. A ,). Даже при интенсивностях стимула, которые в 10 раз превышали пороговые уровни (до 500 мкА), активность ограничивалась первоначально активированными областями, и дальнейшего пространственного распространения не наблюдалось ( n = 9).
Коробчатые диаграммы, изображающие пространственное распространение активности после таламической стимуляции в различных экспериментальных условиях. Пространственное распространение выражается как общая площадь поверхности, на которой были зарегистрированы статистически значимые оптические отклики.
NMDA рецепторы опосредуют распространение активности
Внутрикортикальные синаптические взаимодействия в бочкообразной коре в значительной степени зависят от активации глутаматергических рецепторов NMDA (Gil and Amitai, 1996b; Thomson and Deuchars, 1997; Huang et al., 1998). Поэтому мы пришли к выводу, что активация этих рецепторов может усиливать внутрикортикальные синаптические взаимодействия и увеличивать пространственное распространение активности в бочкообразной коре головного мозга. Чтобы проверить эту гипотезу, мы обрабатывали срезы в растворе, номинально не содержащем магния (0 Mg 2+ ), состояние, которое усиливает активацию рецепторов NMDA (Collingridge and Bliss, 1995).
В 0 Mg 2+ таламическая стимуляция с интенсивностью чуть выше пороговой привела к резким изменениям в пространственно-временных паттернах активации.Как и в контрольных условиях, сначала регистрировались оптические сигналы от одного или двух бочек и границы слоя V – VI непосредственно под ними. Однако активность затем распространяется вертикально на супрагранулярный и инфрагранулярный слои и через границу слоя Vb – VI, причем пики активности в этих областях происходят через 25–45 мсек после стимула (Figs. B ,). Затем активность распространилась на соседние стволы и супрагранулярные слои непосредственно над ними (через 40–64 мсек после стимула) и, наконец, на поверхностный слой II и слой I (через 50–82 мсек после стимула).Аналогичные пространственно-временные паттерны были зарегистрированы для всех срезов, испытанных в растворах 0 Mg 2+ ( n = 17).
Мы оценили скорость внутрикортикального распространения оптических сигналов, измерив задержку между началом оптических откликов в двух точках среза и разделив это значение на расстояние между этими точками. Распространение оптической активности происходило со скоростью 0,112 ± 0,028 м / сек ( n = 17). Поскольку наш расчет не учитывает вклад синаптических задержек, и поскольку мы не можем определить количество синаптических задержек между двумя измеренными точками, это значение может быть ниже, чем фактическая скорость проводимости внутрикортикальных аксонов.Тем не менее, это значение аналогично вычисленному из измерений распространения потенциалов внутрикортикального поля в неокортексе in vitro (Aroniadou, Keller, 1993) и из оценок скоростей внутрикортикальной проводимости в бочкообразной коре in vivo (Армстронг- Джеймс, 1995).
Относительно длительная инкубация кортикальных срезов в растворах, содержащих низкие концентрации Mg 2+ , может вызвать спонтанную эпилептическую активность (Tsau et al., 1998). В нашем препарате мы не наблюдали пароксизмальной активности ни в оптических ответах, ни в потенциалах поля ( n = 17). Это может быть связано с относительно короткими периодами (≤15 мин), которые мы использовали номинально без Mg 2+ -содержащих растворов.
Усиленное распространение активности было значительно ( p <10 -4 ) подавлено применением антагониста рецептора NMDA AP5 (50 мкм; n = 6; Фиг. C ,). Это открытие подтверждает вывод о том, что активация рецепторов NMDA ответственна за усиленное пространственно-временное распространение оптических ответов в условиях 0 Mg 2+ .В этих условиях потенциал-зависимый блок рецепторов NMDA Mg 2+ снимается (Collingridge and Bliss, 1995), что приводит к активации внутрикортикальных синаптических взаимодействий, опосредованных этими рецепторами.
Аналогичное усиление распространения активности в 0 Mg 2+ было зарегистрировано в ответ на стимуляцию внутренней капсулы в срезах, в которых были разорваны связи между таламусом VB и корой ствола ( n = 5). Это предполагает, что активация рецепторов NMDA в коре головного мозга необходима для усиления распространения активности, и что этот эффект не требует активации рецепторов NMDA в таламусе.
Частотно-зависимая модуляция распространения сигнала
В физиологических условиях зависимые от напряжения рецепторы NMDA могут активироваться последовательностями стимулов (Collingridge and Bliss, 1995). Поэтому мы проверили, приведет ли короткая серия стимулов низкой интенсивности (четыре импульса, 30 мкА, 100 Гц) к активации рецепторов NMDA и усилению внутрикортикального распространения активности. Эти параметры стимула были выбраны для имитации последовательности потенциалов действия, вызванных в таламокортикальных нейронах в ответ на отклонения усов (Hartings and Simons, 1998; см. Обсуждение).
В нормальном ACSF последовательность стимулов вызвала пространственно-временные паттерны ответа, которые были подобны тем, которые вызывались в условиях 0 Mg 2+ (фиг. D ,). Активность зародилась в одном или двух барабанах и граница слоя Vb – VI непосредственно под ними и распространялась как по вертикали, так и по горизонтали на соседние столбики с цилиндрами. Однако, в отличие от условия 0 Mg 2+ , высокочастотная стимуляция не приводила к распространению активности на поверхностную часть слоя II и на слой I (рис. D ). Кроме того, ответы на высокочастотную стимуляцию длились значительно дольше (105–222 мс), предположительно отражая временное суммирование ответов на последовательность стимулов. Подобные пространственно-временные паттерны были записаны в 16 срезах в ответ на эти параметры стимула.
Чтобы определить, опосредовано ли усиленное размножение активацией рецепторов NMDA, мы проверили его чувствительность к антагонисту NMDA AP5 (50 мкм). Во всех случаях ( n = 9) AP5 значительно ( p <10 -4 ) ослаблял распространение ответов на высокочастотную таламическую стимуляцию (рис. E ,). Эти данные свидетельствуют о том, что стимуляция таламуса короткой серией импульсов активирует синаптические взаимодействия с участием рецепторов NMDA и приводит к распространению активности, опосредованной внутрикортикальными связями.
Альтернативно, высокочастотная стимуляция могла задействовать дополнительные таламокортикальные нейроны, и этот усиленный афферентный вход может быть ответственным за расширенные паттерны корковой активности. Если бы это было так, увеличение интенсивности стимула должно дать аналогичный результат.Однако, как описано выше (рис. A ), даже при высокой интенсивности стимула одиночный импульс стимула не мог вызвать внутрикортикального распространения активности.
ГАМКергическая модуляция распространения сигнала
ГАМК Ингибирование, опосредованное рецептором , глубоко влияет на свойства ответа нейронов бочкообразной коры (Brumberg et al., 1996; Kyriazi et al., 1996a, b, 1998). Многие из этих свойств ответа опосредуются, по крайней мере частично, внутрикортикальными синаптическими взаимодействиями (Armstrong-James, 1995; Keller, 1995).Чтобы определить роль рецепторов GABA A в модуляции этих внутрикортикальных взаимодействий, мы сравнили кортикальные пространственно-временные модели активности до и после применения бикукуллина, антагониста рецепторов GABA A . Относительно низкие концентрации бикукуллина (1–2 мкм) приводили к увеличению амплитуды оптических ответов (на 50–80%, 63 ± 12%; n = 14), но не влияли на пространственное распространение активности ( Рис. F ). То есть, как и в контрольных условиях, активность была ограничена одним или двумя стволами в слое IV и границей слоя Vb – VI непосредственно под ними; общая площадь, активированная в присутствии бикукуллина, существенно не отличалась от площади, активированной в контрольных условиях ( p <10 -4 ; рис.).
Повышение концентрации бикукуллина до 5 или 10 мкм или инкубация срезов в низкой концентрации бикукуллина в течение> 20 мин приводило к выраженным пароксизмальным (эпилептическим) явлениям, вызванным таламусом. Эти пароксизмальные события отображены в оптической записи, полученной с одного фотодиода, на Рисунке G . Пространственно-временной анализ оптических откликов показывает закономерности распространения этих событий (Рис. H ). Сходные паттерны пароксизмальной активности, вызванной таламом, были зарегистрированы для всех тестируемых срезов ( n = 13).Как и в контрольных условиях, начальная активация происходила в стволе слоя IV и в слоях Vb и VI под ним (рис. H , G, стрелка ). После периода переменной задержки (10–150 мсек) ответ с большой амплитудой исходил от активированного ствола и распространялся на супрагранулярные слои в пределах того же столбца ствола, где он достиг своего пика через 50–130 мсек после стимула. Затем активность распространялась вертикально вдоль того же столбца на инфрагранулярные слои. Затем активность распространялась горизонтально через инфрагранулярный и надгранулярный слои с одинаковой скоростью.Однако пароксизмальная активность в инфрагранулярных слоях сохранялась дольше, чем в супрагранулярных слоях (рис. H ).
И в инфрагранулярном, и в надгранулярном слоях пароксизмальная активность распространялась со скоростью 0,04 ± 0,10 м / сек ( n = 13), что несколько ниже, чем сообщалось Chervin et al. (1988) (∼0.06–0.09 м / с). Это расхождение может быть связано с разницей в температуре записи (34 ° C в исследовании Chervin et al., 1988, по сравнению с комнатной температурой в нашем).Интересно, что относительно небольшая активность была обнаружена в барабанах слоя IV, что привело к появлению двух полос распространяющихся волн, одна в супрагранулярных, а другая в инфрагранулярных слоях (рис. H ).
В некоторых срезах ( n = 5 из 13) вторая пароксизмальная волна появилась через 70–250 мсек после спада первой волны (рис. G, H ). Во всех случаях вторая волна зародилась в инфрагранулярных слоях и распространилась на супрагранулярные слои.Затем активность распространялась горизонтально как через надгранулярный, так и через инфрагранулярный слои, с пространственно-временными характеристиками, аналогичными характеристикам исходных пароксизмальных волн.
Аналогичная пароксизмальная активность была зарегистрирована в ответ на стимуляцию внутренней капсулы в срезах, в которых были разорваны связи между таламусом VB и корой ствола ( n = 3). Это предполагает, что в соответствии с предыдущими исследованиями (Connors, 1984; Chervin et al., 1988; Telfeian and Connors, 1998; Tsau et al., 1998), подавление рецепторов GABA A в коре ствола достаточно, чтобы вызвать пароксизмальную активность.
ОБСУЖДЕНИЕ
Технические соображения
Способность идентифицировать отдельные функциональные столбцы в неокрашенном препарате in vitro позволила нам изучить распространение таламических входов через внутриколоночные и межколоночные кортикальные пути. Парадигма функциональной визуализации, которую мы использовали, выгодна тем, что позволяет анализировать с высоким разрешением пространственно-временное распространение активности в пределах коры бочонка и модуляцию этой активности с помощью различных манипуляций.Оптические сигналы, зарегистрированные в нашем исследовании, особенно чувствительны к сверхпороговой (то есть пиковой) активности в синхронно активированной популяции нейронов. Это связано с тем, что оптические отклики, связанные с красителями, имеют относительно низкое отношение сигнал / шум (Wu and Cohen, 1993). Чувствительность наших записей дополнительно ограничивается использованием объективов с относительно низким увеличением, которые необходимы для одновременной записи активности из больших областей в срезе. Поэтому возможно, что в наших условиях записи подпороговые или надпороговые ответы отдельных нейронов не были обнаружены.
Активность, вызванная таламусом
В ответ на низкоинтенсивную стимуляцию таламуса VB активность была ограничена одним или двумя барабанами. Это говорит о том, что стимуляция активировала нейроны в соответствующем количестве таламических баррелоидов, потому что таламокортикальные нейроны в пределах одного баррелоидного проекта предпочтительнее одного кортикального ствола (Killackey and Leshin, 1975; Land et al., 1995; Keller and Carlson, 1999). Увеличение интенсивности стимула приводило к одновременной активации дополнительных стволов, предположительно отражая рекрутирование таламокортикальных нейронов в соседние таламические баррелоиды.
Ответы, зарегистрированные из стволов слоя IV, скорее всего, представляют постсинаптическую активацию как возбуждающих, так и тормозных стволовых нейронов, поскольку обе популяции получают относительно большое количество таламокортикальных синапсов (Benshalom and White, 1986; White, 1986) и отвечают моносинаптически на таламическую стимуляцию ( Swadlow et al., 1998; Gil et al., 1999; Katz et al., 1999). Оптические ответы в стволах могут также происходить из сегментов слоя IV дендритов, принадлежащих клеткам, чьи родительские соматы находятся в других корковых слоях, которые, как известно, получают таламокортикальные синапсы (White, 1986).Они могут включать пирамидные нейроны слоев V и VI, которые способны продуцировать дендритные шипы, опосредованные Na + — и Ca 2+ , в ответ на синаптические входы (Kim and Connors, 1993; Markram and Sakmann, 1994; Yuste et al. др., 1994; Schwindt, Crill, 1997).
В дополнение к ответам в слое IV, таламическая стимуляция также вызывала оптические сигналы около границы слоя Vb – VI непосредственно под активированными стволами. Эти ответы, скорее всего, исходят от сомов нейронов, которые получают мощные таламические входы, либо от таламокортикальных окончаний в слое IV, либо от меньшего количества таламических окончаний в инфрагранулярных слоях (Keller et al., 1985; Chmielowska et al., 1989). Эти клетки, вероятно, включают кортикоталамические клетки, которые получают относительно высокую плотность таламических синапсов на своих апикальных дендритах в слое IV (White and Hersch, 1982; Keller and White, 1989). В большинстве случаев ответы в слое Vb – VI происходили одновременно или даже раньше, чем ответы в слое IV. Это согласуется с данными регистрации отдельных клеток, демонстрирующими, что нейроны в инфрагранулярных слоях реагируют на отклонения усов одновременно с их аналогами в слое IV (Simons, 1978; Chapin, 1986; Swadlow, 1989; Armstrong-James et al., 1992).
Это открытие предполагает, что инфрагранулярные нейроны, вероятно, участвуют на ранних стадиях обработки входных данных усов. Действительно, поскольку коллатерали аксонов многих инфрагранулярных нейронов проецируются в слой IV (Staiger et al., 1996; Gottlieb and Keller, 1997; Zhang and Deschênes, 1997), эти клетки могут играть роль в определении свойств клеток слоя IV. — традиционно рассматриваемые как начальные участки обработки таламокортикальной информации. Таким образом, классический взгляд на то, что таламический вход в кору бочонка (и другие области коры, например.g., Eccles, 1984) сначала «обрабатываются» клетками слоя IV, и то, что инфрагранулярные нейроны, которые проецируются на подкорковые мишени, составляют конечную «выходную» стадию корковой обработки, можно упростить.
Потенциальным осложнением является возможность того, что таламическая стимуляция также антидромно активировала кортикоталамические нейроны и вызвала ответы в кортикальных нейронах, постсинаптических по отношению к коллатералям аксонов этих клеток. Это маловероятно, потому что порог стимула для активации таламокортикальных (TC) аксонов примерно на 1 порядок ниже, чем ток, необходимый для антидромной активации кортикоталамических (CT) клеток (Ferster and Lindström, 1985; Stratford et al., 1996; Swadlow, 1998). Кроме того, в предыдущих исследованиях сообщалось, что пучки аксонов TC и CT перемещаются в разных переднезадних плоскостях во внутренней капсуле, так что срезы, содержащие афференты TC, редко включают аксоны CT (Agmon and Connors, 1991; Agmon and Connors, 1992). Наконец, ожидается, что антидромная стимуляция вызовет оптические сигналы в соматах CT, и эти сигналы должны быть устойчивыми к подавлению синаптической передачи; однако все оптические отклики подавлялись в условиях 0 Ca 2+ (рис.).
GABA
A рецептор-опосредованные ответыОтносительно низкие концентрации бикукуллина антагониста рецептора GABA A привели к выраженному увеличению амплитуды таламических ответов, но не увеличили пространственное распространение этих ответов. Это открытие предполагает, что ГАМКергическое ингибирование не оказывает значительного влияния на активацию внутриколоночных или межколоночных путей в коре ствола.
Применение антагонистов рецептора GABA A in vivo приводит к увеличению размера окружающего рецептивного поля бочкообразных нейронов слоя IV, так что ответы этих клеток на неглавные усы резко усиливаются (Brumberg et al., 1996; Кириази и др., 1996b). Наше открытие, что низкие концентрации бикукуллина не оказывают значительного влияния на межколоночные синаптические взаимодействия, предполагает, что наблюдаемые расширения рецептивного поля in vivo связаны с модуляцией синаптических взаимодействий внутри отдельных стволов, а , а не , зависят от изменений межколоночных взаимодействий. Это дополнительно подтверждает гипотезу, сформулированную Саймонсом и соавторами (Simons and Carvell, 1989; Simons, 1995; Goldreich et al., 1999), что свойства ответа стволовых нейронов слоя IV формируются главным образом за счет тормозных и возбуждающих синаптических взаимодействий, ограниченных одним стволом.
В соответствии с предыдущими исследованиями (Gil and Amitai, 1995; Telfeian and Connors, 1998), в присутствии более высоких концентраций бикукуллина таламическая стимуляция приводила к пароксизмальной или эпилептической активности в коре головного мозга. Считается, что пространственное распространение пароксизмальной активности происходит в первую очередь за счет активации внутрикортикальных возбуждающих синаптических путей (Gutnick et al., 1982; Трауб и Вонг, 1982; Червин и др., 1988; Коннорс и Амитаи, 1997). Следовательно, анализ этой активности предоставляет полезный инструмент для выяснения внутрикортикальных синаптических путей (Fleidervish et al., 1998).
Вызванная таламом пароксизмальная активность была инициирована в цилиндре слоя IV и сначала распространилась на супрагранулярные слои непосредственно над этим цилиндром. Это согласуется с сообщениями об инициировании и вертикальном распространении электрически записанных эпилептических разрядов в неокортексе (Connors, 1984).Анатомическими субстратами для распространения к супрагранулярным слоям, скорее всего, являются вертикальные коллатерали аксонов непирамидных нейронов слоя IV (Simons and Woolsey, 1984; Katz et al., 1999). Последующее распространение активности на инфрагранулярные слои, вероятно, опосредуется нисходящими коллатералями аксонов, принадлежащими супрагранулярным пирамидным клеткам (Bernardo et al., 1990; Gottlieb and Keller, 1997; Katz et al., 1999). Горизонтальное интракортикальное распространение пароксизмальной активности опосредуется внутрикортикальными синаптическими взаимодействиями (Connors and Amitai, 1993), наиболее вероятно с участием коллатералей аксонов супрагранулярных и инфрагранулярных пирамидных клеток (Bernardo et al., 1990; Готтлиб и Келлер, 1997). В соответствии с предыдущими исследованиями, скорость этого распространения была значительно ниже, чем скорость проводимости внутрикортикальных аксонов (Gutnick et al., 1982; Telfeian and Connors, 1998). Это открытие подтверждает гипотезу о том, что эпилептическая активность распространяется за счет синхронизации активности ограниченных корковых цепей (Traub and Wong, 1982).
Хотя пароксизмальная активность большой амплитуды распространялась через надгранулярный и инфрагранулярный слои, эти распространяющиеся волны в основном избегали стволов слоя IV (рис. H ). Этот вывод согласуется с данными, предполагающими, что существует относительно мало прямых связей между полостями соседних стволов (Simons, Woolsey, 1984; Goldreich et al., 1999; Katz et al., 1999; Kim and Ebner, 1999). Этот результат также согласуется с данными, демонстрирующими, что, хотя пароксизмальная активность может инициироваться в слое IV (Connors, 1984), она может распространяться горизонтально через супрагранулярные или инфрагранулярные слои, даже если эти слои хирургически изолированы от слоя IV (Telfeian and Connors , 1998).
Ответы, опосредованные рецептором NMDA
В отличие от ограниченных эффектов низких концентраций бикукуллина, активация рецепторов NMDA привела к резкому распространению вызванной таламиком активности по всей коре головного мозга. Эти находки предполагают, что рецепторы NMDA критически вовлечены во внутрикортикальные синаптические взаимодействия в бочкообразной коре, что согласуется с предыдущими данными in vitro (Thomson et al., 1988; Gil and Amitai, 1996a). Действительно, наши данные предполагают, что надпороговое внутрикортикальное распространение таламической вызванной активности требует активации рецепторов NMDA.Это согласуется с данными in vivo о том, что надпороговые, межколоночные синаптические взаимодействия, по крайней мере, в надгранулярных слоях бочкообразной коры, полностью зависят от активации рецепторов NMDA (Huang et al., 1998) и что свойства ответа нейронов в соматосенсорной коре сильно влияют манипуляции с рецепторами NMDA (Armstrong-James et al., 1993; Whitsel et al., 1999).
Мы еще не идентифицировали нейроны, ответственные за опосредование надпороговой, зависимой от рецептора NMDA распространения активности.Хотя таламокортикальные синапсы активируют как NMDA-рецепторы, так и не-NMDA-рецепторы (Gil and Amitai, 1996a), большая эффективность этих синапсов (Gil et al., 1999) предполагает, что они вряд ли будут дополнительно усилены за счет разоблачения NMDA-рецепторов. Напротив, значительно меньшая эффективность внутрикортикальных синаптических взаимодействий (Gil et al., 1999) делает их особенно чувствительными к такому усилению. В самом деле, наши находки подтверждают, что активность, зависимая от рецептора NMDA, распространяется через внутрикортикальные связи от нейронов-реципиентов таламуса в слоях IV и на границе V-VI к их постсинаптическим мишеням в тех же и соседних столбцах ствола.
Ответы на повторяющуюся стимуляцию
Помимо своей роли в развитии синапсов и пластичности, рецепторы NMDA играют важную роль в качестве детекторов совпадений (Collingridge and Singer, 1990; Markram et al., 1997). То есть зависимость этих рецепторов от напряжения делает их особенно чувствительными к синаптическим входам, которые происходят, когда постсинаптическая клетка деполяризуется, например, более ранними входами в повторяющейся последовательности. Здесь мы сообщаем, что повторяющаяся стимуляция таламокортикальных афферентов приводит к усиленному распространению активности в бочкообразной коре головного мозга за счет активации зависимых от рецептора NMDA внутрикортикальных путей.
Во время тактильного различения крысы метаются на частотах от 1 до 20 Гц, с доминирующей частотой ~ 8 Гц (Carvell and Simons, 1990; Kleinfeld et al., 1999). Таламокортикальные нейроны могут быть захвачены отклонениями усов с частотой не менее 12 Гц (Hartings and Simons, 1998) и реагируют на одиночных отклонений усов, в среднем, двумя спайками с частотой ~ 350 Гц (Kyriazi et al., 1994). Таким образом, параметры стимула, проверенные в настоящем исследовании (четыре импульса с частотой 100 Гц), находятся в пределах физиологических параметров, которые могут иметь место во время исследовательского взбивания.
Почти все исследования in vivo бочковой коры проверяли свойства рецептивного поля нейронов бочкообразной коры в ответ на одиночные отклонения усов. Наши результаты показывают, что во время высокочастотного исследовательского взбивания реакции этих нейронов будут кардинально отличаться. В частности, из-за частотно-зависимой активации внутрикортикальных связей, опосредованных рецептором NMDA, ответы нейронов бочкообразной коры на неглавные усы (окружающее рецептивное поле) будут значительно больше, чем те, которые регистрируются в ответ на одиночные отклонения усов.
Экономические опасения снижают цены на нефть на второй день
Но, хотя спрос в США и Западной Европе падает, потребление нефти в этом году, как ожидается, вырастет из-за роста в Китае, Индии и на Ближнем Востоке. Этот рост по-прежнему является основанием для цен на нефть.
Падение цен усугубилось ослаблением геополитической напряженности, поддерживавшей успехи прошлой недели, особенно вокруг Ирана, второго по величине производителя нефти ОПЕК, а также более позитивными новостями из Нигерии, где добыча возобновилась после недавних перебоев.
Джон Килдафф, энергетический аналитик брокерской фирмы MF Global, сказал, что новости о прямых переговорах между Соединенными Штатами и Ираном и неожиданном увеличении запасов нефти были «одним-двумя ударами по медведям», которые могут сигнализировать о более крутом падение цен в ближайшие недели.
С 2002 года цены на нефть выросли более чем в семь раз, поскольку быстрорастущий спрос в развивающихся странах опередил новые поставки нефти. Это привело к тому, что в Конгрессе были предприняты усилия по сокращению чрезмерных затрат на энергию, например, путем увеличения объемов морского бурения для увеличения внутренних поставок.
Согласно опросу MasterCard, спрос на бензин в США упал на 5,2 процента на прошлой неделе, это 12-е недельное падение подряд. С начала года спрос на бензин упал примерно на 1 процент по сравнению с тем же периодом прошлого года.
В результате повышения цен на газ и снижения спроса нефтеперерабатывающие предприятия используют меньше нефти. Вместо того, чтобы падать по мере того, как нефтепереработчики используют свои запасы, как это обычно бывает в это время года, запасы нефти наращиваются. Запасы нефти выросли 2.На прошлой неделе министерство энергетики сообщило, что на прошлой неделе было 95 миллионов баррелей, или 296,9 миллиона баррелей. Аналитики ожидали, что запасы упадут примерно на 2,2 миллиона баррелей.
Через день после того, как в своих показаниях в Сенате г-н Бернанке дал в целом мрачную оценку экономики, он попытался заверить членов комитета Палаты представителей в том, что экономика выздоровеет.
«Мы преодолеем эти финансовые бури», — сказал он собеседнику. «Мы будем работать над этим циклическим движением, которое у нас есть.И экономика вернется к хорошему росту. Но нам нужно поработать над несколькими вещами, чтобы добиться этого ».
Границы | Динамика индуцированного гипоксией отека тканей в коре ствола крысы in vitro
Введение
Отек мозга — серьезное опасное для жизни осложнение ишемического поражения головного мозга (Stokum et al., 2016; Zheng et al., 2016). Отек мозга вызывает масс-эффект с деформацией, сдвигом тканей и повышенным внутричерепным давлением, что приводит к церебральной грыже, дальнейшему повреждению мозга и смерти (Frank, 1995; Schwab et al., 1996; Достович и др., 2016). Однако эффективное лечение отеков у пациентов с инсультом, включая гипервентиляцию, гиперосмотическую терапию, диуретики, кортикостероиды или барбитураты, ограничено. В тяжелых случаях отека требуется хирургическая декомпрессия, чтобы создать пространство для размещения увеличенного объема, созданного опухшим мозгом, путем открытия свода черепа и твердой мозговой оболочки или даже путем удаления нежизнеспособной или несущественной ткани мозга (Dostovic et al., 2016; Zheng и др., 2016).
Отек мозга вызывается увеличением содержания воды в тканях мозга в результате осмотических изменений, вызванных нарушениями ионного градиента, и накоплением осмотически активных метаболитов, таких как лактат, в ишемической области (Stokum et al., 2016). Отек развивается в несколько стадий, начиная с цитотоксического отека или клеточного набухания, проявляющегося через несколько минут после метаболического инсульта в результате транслокации воды из внеклеточного во внутриклеточный компартмент, вызванного нарушением трансмембранных ионных градиентов во время аноксической распространяющейся деполяризации (ASD) (Dreier и др., 2018). Цитотоксический отек начинает развиваться во время АСД и характеризуется набуханием нейронов и глиальных клеток, сужением дендритов, сокращением внеклеточного пространства и изменениями оптической прозрачности ткани (Vanharreveld, 1957; Murphy et al., 2008; Ришер и др., 2009, 2010, 2012). Подобные явления также наблюдаются в погруженных кортикальных срезах, подвергшихся кислородно-глюкозной депривации (OGD) (Obeidat et al., 2000; Andrew et al., 2007; Risher et al., 2009; Brisson and Andrew, 2012).
Несмотря на обширные знания о динамике набухания клеток, начало которой тесно связано с АСД, динамика набухания тканей менее изучена. Важно отметить, что набухание клеток может происходить без набухания тканей. Если доступ воды к ишемической ткани ограничен, набухание клеток происходит за счет перемещения воды из внеклеточного во внутриклеточное пространство без каких-либо изменений в общем объеме ткани мозга.Например, при ишемии мозга, вызванной остановкой сердца, общий объем мозговой ткани и содержание в ней воды не изменяются. Точно так же ишемическое ядро не увеличивается в размере, несмотря на цитотоксическое набухание, описанное в Klatzo (1987) и Stokum et al. (2016). Однако значительный отек ткани может возникнуть, если доступен внешний доступ воды к ишемизированной ткани, например, в препаратах погруженных срезов, где срез подвергается воздействию электролитов и воды в среде для суперфузии ad libitum .В самом деле, содержание воды увеличивается в срезах головного мозга, подверженных ишемическим состояниям, включая OGD и другие метаболические нарушения (Kumar et al., 2006; Elkin et al., 2010). Соответственно, увеличение объема среза после комбинации отравления цианидом с агликемией можно было обнаружить уже через 30 минут после метаболического инсульта (Elkin et al., 2010). Кроме того, предыдущие исследования показали технические проблемы с покадровой визуализацией срезов мозга, вызванные сдвигами фокальной плоскости во время OGD, что требовало взятия образцов оптического сечения на лету, а также повторного центрирования и регулировки поля фокуса до получения изображения. стеки (Эндрю и др., 2007; Ришер и др., 2009). Подобная техническая проблема была указана в записях патч-клампа из интактного гиппокампа in vitro , где электрический доступ к регистрируемым нейронам часто терялся после индуцированного OGD aSD, вероятно, из-за движения ткани (Dzhala et al., 2001). Однако динамика ишемического движения тканей головного мозга и отека в срезах головного мозга ранее не рассматривалась.
В настоящем исследовании мы исследовали динамику движения ткани и объемные изменения, происходящие в срезах коры ствола крысы во время ишемических состояний OGD.Мы обнаружили, что OGD эффективно вызывает движение массы и набухание тканей, которые характерны для ишемического отека мозга, что начало набухания ткани тесно связано с aSD и что оно развивается в относительно медленном временном масштабе после aSD.
Материалы и методы
Этическое разрешение
Все протоколы использования животных соответствовали рекомендациям Национального института здравоохранения и медицинских исследований Франции (INSERM, протокол N007.08.01) и Казанского федерального университета по использованию лабораторных животных (этическое одобрение Институционального комитета по уходу и использованию животных Казанский государственный медицинский университет №9-2013).
Подготовка ломтиков мозга
Использоваликрыс линии Вистар (возраст 16–23 дня) обоего пола. Животных декапитировали под анестезией изофлураном (5%), мозг быстро извлекали и помещали в ледяной (2–5 ° C) раствор для срезов [модифицировано из (Dugue et al., 2005)] следующего состава (в мМ): K-глюконат 140, Na-глюконат 15, NaCl 4, EGTA 0,2, D-AP5 50 мкМ и HEPES 10 (pH 7,4). Таламокортикальные срезы толщиной 400 мкм вырезали с использованием вибратома PELCO easiSlicer ™ (Ted Pella, Inc., Реддинг, Калифорния, США). Срезы, содержащие бочкообразную кору, отбирались по анатомическим координатам (Хазипов и др., 2015) и наличию бочкообразных структур в L4. Сначала срезы хранили в насыщенной кислородом (95% O 2 -5% CO 2 ) искусственной спинномозговой жидкости (ACSF) следующего состава (в мМ): NaCl 126, KCl 3,5, CaCl 2 2, MgCl 2 1,3, NaHCO 3 25, NaH 2 PO 4 1,2 и глюкоза 11 (pH 7,4) в течение 30 мин при 32 o C, а затем при комнатной температуре (20–22 ° C) в течение минимум за 1 час до использования.Для записи срезы помещали в погружную камеру и заливали кислородом ACSF при 30–32 ° C со скоростью потока 10 мл / мин. Срезы удерживались на месте в записывающей камере с помощью U-образной рамы ручной работы из серебряной проволоки (0,025 дюйма, A-M Systems, США). Чтобы ткань могла смещаться во время отека, рамка удерживала только две дальние границы среза. Депривация кислорода / глюкозы (OGD) вызывалась суперфузией с ACSF, в которой N 2 заменял O 2 , а сахароза заменяла глюкозу в эквимолярной концентрации.
Электрофизиологические записи
Внеклеточные записи потенциалов локального поля (LFP) были выполнены в коре головного мозга с использованием односайтовых стеклянных пипеточных электродов. Электроды вытягивались из капилляров из боросиликатного стекла (BF150-86-10, Sutter Instrument, Novato, CA, USA) и имели сопротивление 2–3 МОм при заполнении ACSF. Электроды подсоединяли через хлорированную серебряную проволоку к головному блоку усилителя с патч-зажимом MultiClamp700B (Axon Instruments, Юнион-Сити, Калифорния, США).Запись производилась в режиме фиксированного постоянного напряжения, затем инвертировались токи и калибровалось напряжение с шагом 5 мВ.
Интервальная съемка
Запись с помощью покадровой микроскопии выполнялась с использованием вертикального микроскопа BX51WI, оснащенного сухим объективом 4x / 0,10 Plan N и погруженным в воду объективом 40x / 0,80 LUMPlanFL N (Olympus, Токио, Япония). Срез освещали галогенной лампой с полосовым фильтром 775 нм. Изображения были получены с помощью камеры QIClick-R-F-M-12 CCD (QImaging, Surrey, BC, Canada) с разрешением 696 × 520 или 1392 × 1040 пикселей и скоростью сбора данных 5–10 кадров / с.
Покадровая съемка кортикальной поверхности с поправкой на изменение высоты поверхности выполнялась либо путем фокусировки на заполненных флуоресцентным красителем клеток вручную, либо путем автофокусировки на изображениях поверхности срезов с самым высоким контрастом с использованием самодельной Z-коррекции. система. Отдельные нейроны слоя 4, расположенные близко к поверхности среза, метили флуоресцентным красителем CF ™ 488A (100 мкМ) через патч-пипетку в режиме целых клеток. Затем пипетку удаляли, чтобы обеспечить свободное перемещение клеток.Система автофокусировки состояла из шагового двигателя, который был прикреплен к Z-приводу микроскопа и управлялся платой управления Arduino Uno, и программного обеспечения LabVIEW (National Instruments) с модулем Vision Development для определения фокальной плоскости самого высокого контрастное изображение посредством онлайн-анализа нескольких изображений, сделанных на разных расстояниях фокусировки (диапазон: от +50 до -100 мкм, с шагом 4 мкм). Размер изображения был изменен до разрешения 696 × 520 пикселей. Затем оператор Собеля использовался для определения градиента яркости.Статистическое распределение пикселей разной яркости рассчитывалось после оператора Собеля. Средняя яркость в данном распределении была критерием резкости изображения. Один цикл автофокусировки длился в среднем 6,6 ± 0,9 с. Дальнейший автономный анализ положения объектов на корковой поверхности был использован для обнаружения дрейфа ткани в горизонтальной плоскости.
Анализ LFP и OIS
Данные были проанализированы с использованием специально написанных процедур в Matlab (MathWorks, Inc., Натик, Массачусетс, США). OIS рассчитывалась методом вычитания первого кадра: OIS (t) = (I (t) — I 0 ) / I 0 , где I (t) — интенсивность пикселей в момент t, I 0 — усредненная по времени интенсивность пикселей в предварительно подготовленном базовом периоде (100 с). Полученные кадры были отфильтрованы медианным фильтром 10 × 10. Области интереса (ROI) были выбраны как квадратные области рядом с местом записи. Трассы OIS были рассчитаны как средний сигнал OIS в выбранных областях интереса.
сигналов LFP были субдискретизированы до 1 кГц.Подгонка непрерывной бегущей строки была удалена с помощью локальной линейной регрессии в окнах 300 с с перекрытием 10 с [ функция locdetrend из набора инструментов Chronux (http://chronux.org/)]. Данные были сглажены фильтром скользящего среднего из 1000 точек, и был обнаружен локальный отрицательный пик aSD. Базовый уровень рассчитывали как среднее значение LFP во временном окне от -20 до -10 с, предшествующем SD. Амплитуда aSD рассчитывалась как максимальный отрицательный пик LFP от базовой линии.
Статистический анализ
Статистический анализ был основан на непараметрическом тесте суммы рангов Уилкоксона (парные выборки) с уровнем значимости, установленным на p <0.05. Результаты представлены в виде медианы ± межквартильный размах, где межквартильный размах - это разница между 75-м и 25-м процентилями.
Результаты
Мы исследовали динамику тканевых сдвигов и объемных изменений в срезах бочкообразной коры головного мозга крысы во время ишемических состояний, индуцированных OGD. С этой целью мы использовали покадровую съемку поверхности среза и границ с сопутствующим потенциалом местного поля и записи OIS. В соответствии с результатами предыдущего исследования с использованием этого препарата (Juzekaeva et al., 2017), OGD индуцировал aSD с задержкой 9,7 ± 3,2 мин ( n = 54 среза) после начала перфузии раствором OGD. aSD достигло пикового отрицательного значения 5,8 ± 3,9 мВ в L4 ( n = 54), и это также было связано с резким переходным увеличением прозрачности среза на 25,7 ± 23,2% dI / I ( n = 49) (рисунки 1B, C; см. Также сводный рисунок 6).
Рисунок 1 . Кислородно-глюкозная депривация (OGD) индуцировала движение массы ткани: общее наблюдение. (A) Моментальные снимки поверхности кортикального среза в различные моменты времени из вызванной OGD аноксической распределяющей деполяризации (ASD), полученные при постоянной фокальной плоскости, установленной на поверхности среза до OGD. Обратите внимание, что ткань начинает дрейфовать после SD-карты и смещается из фокальной плоскости; (a – h) в верхнем левом углу указывают моменты времени на графике B, когда были сделаны снимки. Обратите внимание на изменения внешнего вида сосуда (v) из-за вертикального смещения ткани. (B, C) Соответствующие следы LFP (B) и OIS (C) записи из кортикального ствола.Интересующая область (ROI) записи OIS обозначена пунктирной рамкой на снимке (a). См. Также соответствующее онлайн-видео 1.
Массовое движение тканей: общее наблюдение
При первоначальной покадровой визуализации кортикальной поверхности с постоянной фокальной плоскостью (рис. 1A и видео 1) мы наблюдали, что во время перфузии среза раствором OGD вся кортикальная ткань начала дрейфовать в горизонтальной плоскости, о чем свидетельствует смещение различных объектов, таких как клетки и сосуды, а также по вертикали, о чем свидетельствует смещение объектов из фокальной плоскости и появление новых движущихся объектов (см. также Видео 2 при малом увеличении, чтобы узнать об исчезновении сосудов).Изображения также стали размытыми, теряли контрастность объектов. Особенно впечатляющим было движение ткани относительно неподвижных объектов, таких как пипетка для регистрации потенциала внеклеточного поля (видео 1). Эти тканевые дрейфы начинались в связи с aSD и продолжались после aSD (Рисунки 1B, C). Мы также попытались количественно оценить индуцированное OGD смещение тканей в нескольких настройках визуализации.
Расширение границ среза
Сначала мы охарактеризовали сдвиги границ срезов.С этой целью мы проанализировали положение верхней (менингеальной) и нижней (желудочковой) границ кортикального среза в области коры соматосенсорной бочки. Как показано на фиг. 2A – C и видео 2, границы срезов оставались неподвижными во время контрольных условий, на ранней стадии OGD и индуцированного OGD aSD. Однако сразу после ASD границы срезов начали расширяться, и этот процесс продолжался после реперфузии срезов оксигенированной глюкозосодержащей ACSF. Поскольку начало расширения границы, а также другие объемные изменения, вызванные OGD (см. Ниже), были тесно связаны с aSD, а не с длительностью OGD, анализ групповых данных проводился в отношении aSD с пиком отрицательности aSD, взятым как контрольный момент времени T = 0.Границы начали смещаться из контрольного положения в среднем через 3,5 ± 1,8 мин ( n = 21) после aSD (рис. 2E). Через 29 ± 23 мин после АСД (период, который включал 15,1 ± 4,5 мин при ОГД и 8,6 ± 4,1 мин реперфузии), верхняя (менингеальная) и нижняя (желудочковая) границы расширились на 87 ± 44 и 44 ± 68 мкм от их исходные положения до OGD, соответственно ( n = 21). Соответствующая ширина среза (расстояние между верхней и нижней границами) увеличилась на 132 ± 76 мкм, то есть до 107 ± 3% от контрольных значений ( n = 21; рисунки 2D, F).
Рисунок 2 . OGD-индуцированное расширение границ кортикального среза. (A) Микрофотография кортикального среза. Области интереса с менингеальной (синяя рамка) и желудочковой (красная рамка) границами столбца кортикального ствола. (B) Снимки положения верхней менингеальной (синяя линия) и нижней желудочковой (красная линия) границ кортикального среза в пределах ROI на панели A в контроле и в различные моменты времени из индуцированного OGD ASD. Черные линии показывают положение границ до OGD.Обратите внимание, что границы среза начинают расширяться сразу после SD и продолжают расширяться после реперфузии среза с оксигенированной глюкозой, содержащей ACSF. (C) Динамика изменения верхней (синей) и нижней (красной) границ индуцированного OGD смещения из контрольного положения, относящегося к aSD, как нулевой момент времени в эксперименте, показанном на панелях (A, B) . Общий (черный) сдвиг границы — это сумма сдвига верхней и нижней границы. Обратите внимание, что смещение верхней границы более важно, чем смещение нижней границы. (D) Среднее изменение общего расстояния между верхней и нижней границами от n = 21 срез (заштрихованная область показывает SD). Данные относятся к времени aSD = 0. Вертикальные границы на сером поле OGD над графиком указывают среднее начало и смещение OGD от aSD, а горизонтальные линии указывают SD. (E, F) Гистограммы начала расширения границ (E) и полного расширения границ среза (F) 30 мин после SD от n = 21 срез.Движение границ во время эксперимента, показанного на (A – C) , также представлено в онлайн-видео 2.
Высота поверхности среза
Затем мы количественно оценили дрейф массы ткани, вызванный OGD, в середине среза на уровне кортикальных стволов (слой 4). Для этого мы использовали два подхода. В первом подходе для оценки смещения ткани использовалась ручная фокусировка на ячейке, заполненной флуоресцентным красителем. Как показано на рисунке 3A, помеченный нейрон начал дрейфовать в горизонтальной (x-y) плоскости вскоре после SD, а затем исчез из фокальной плоскости, указывая на вертикальное (z) смещение.Ручная коррекция фокальной плоскости выявила смещение клетки вверх от исходного положения (рис. 3C). Первое вертикальное смещение на 14 ± 3 мкм было обнаружено через 7,0 ± 1,8 мин после SD ( n = 7 ячеек). Непрерывный ручной мониторинг высоты фокальной плоскости ячейки выявил дальнейшее прогрессивное смещение ячейки вверх и по горизонтали. В большинстве записей клетки были смещены из оптического поля, что также требовало корректировки координат x-y оптического поля.Восстановленная траектория клетки в трехмерных координатах показана на рисунке 3B (см. Также видео 3). В среднем, клетки дрейфовали после ASD, чтобы достичь своего окончательного смещения 42 ± 7 и 36 ± 5 мкм ( n = 7 клеток) от положения до OGD через 30 минут после aSD в вертикальной и горизонтальной плоскостях, соответственно (Рисунок 3D) .
Рисунок 3 . OGD-индуцированное поднятие поверхности кортикального среза: ручная фокусировка на отдельные клетки. (A) Моментальные снимки нейрона L4, меченного CF ™ 488A, расположенного близко к поверхности кортикального среза в разные моменты времени от индуцированного OGD aSD (точные временные точки от aSD указаны над снимками).aSD в момент времени = 0 обозначается вертикальной стрелкой. Фокальная плоскость регулировалась вручную в ходе эксперимента, и соответствующее вертикальное смещение объектива указано на Z-шкале. Обратите внимание, что после SD-карты ячейка перемещается вверх и по горизонтали. (B) Соответствующая 3D-траектория смещения клеток, вызванного OGD. Время обозначено цветом. (C) Временной ход вертикального смещения ячейки показан на (A, B) . (D) Групповые данные о вертикальном смещении клеток по отношению к индуцированному OGD aSD.Объединенные данные из n = 7 срезов.
Поскольку оценки смещения тканей с использованием ручной коррекции фокальной плоскости имеют ограниченное пространственное и временное разрешение, мы дополнительно оптимизировали определение высоты кортикальной поверхности с помощью самодельной системы автофокусировки, которая позволила настроить фокальную плоскость на изображение с максимальной контрастностью, совпадающее с поверхностью среза, благодаря онлайн-анализу нескольких изображений, сделанных с разным фокусным расстоянием, и обратной связи с электрическим z-приводом.Дальнейший автономный анализ положения объектов на корковой поверхности использовался для обнаружения смещения тканей в горизонтальной плоскости. Покадровая съемка с помощью этой системы автофокусировки дала результаты, аналогичные результатам, полученным при ручной коррекции фокальной плоскости с использованием мониторинга меченых клеток, но с более высоким пространственным и временным разрешением (рисунки 4A – C). Мы обнаружили, что ткань начинает дрейфовать через 1,8 ± 0,8 мин ( n = 4) после aSD, чтобы достичь смещения 71 ± 3 и 26 ± 10 мкм ( n = 4) через 30 мин после aSD в вертикальном и горизонтальном направлениях. , соответственно.Повышение поверхности среза соответствовало увеличению толщины среза до 115 ± 4% ( n = 11) от контрольного значения 400 мкм.
Рисунок 4 . OGD-индуцированное возвышение поверхности кортикального среза: автофокусировка на поверхности среза. (A) Моментальные снимки поверхности среза в различные моменты времени из-за вызванного OGD aSD (точные моменты времени из aSD указаны над снимками). aSD в момент времени = 0 обозначается вертикальной стрелкой. Во время эксперимента фокальная плоскость автоматически настраивалась на высоту максимально контрастного изображения.Соответствующие значения вертикального смещения объектива указаны на Z-шкале. (B) Соответствующая 3D-траектория смещения поверхности среза, вызванного OGD. Время обозначено цветом. (C) График вертикального смещения поверхности среза во времени. См. Соответствующее онлайн-видео 3. (D) Сгруппировать данные о вертикальном смещении поверхности среза по отношению к индуцированному OGD aSD. Объединенные данные из n = 4 среза. Желтые точки указывают на начало вертикального смещения из отдельных экспериментов.Компоновка как на рисунке 2D. См. Также онлайн-видео 3.
Гиперосмотическое лечение
Осмотическая терапия — одно из наиболее часто используемых клинических методов лечения ишемического отека мозга. Поэтому мы дополнительно исследовали, эффективна ли осмотическая терапия в случае отека, вызванного OGD, в срезах коры головного мозга. С этой целью срезы, которые ранее подвергались воздействию OGD и испытывали aSD и набухание, подвергались суперфузии с гиперосмотическим ACSF, содержащим 150 мМ сахарозы, и имели осмолярность 472 мОсм по сравнению с 321 мОсм контрольного ACSF.Как показано на рисунке 5, гиперосмотический раствор вызывал двухфазный ответ с кратковременным увеличением высоты поверхности среза с достижением пика 21 ± 8 мкм ( n = 11) от предсахарозного уровня через 2,2 ± 1,0 мин после сахарозы. нанесение с последующим уменьшением высоты кортикальной поверхности до -35 ± 9 мкм от уровня до сахарозы через 15 мин после обработки гиперосмотическим раствором. Это все еще оставалось выше контрольных уровней до OGD на 35 ± 10 мкм ( n = 11; p <0.05).
Рисунок 5 . Гиперосмотическое лечение уменьшает отек срезов, вызванный OGD. (A) Снимки поверхности срезов в различные моменты времени во время экспериментальной осмотической терапии, проведенной после развития отека, вызванного OGD. Фокальная плоскость была отрегулирована автоматически, как показано на рисунке 4. Обратите внимание, что суперфузия с гиперосмотическим раствором сахарозы уменьшает высоту поверхности среза. (B) Соответствующая 3D-траектория перемещения поверхности среза.Время обозначено цветом. (C) График вертикального смещения поверхности среза во время OGD и после гиперосмотической обработки. (D) Групповые данные о вертикальном смещении поверхности среза по отношению к началу гиперосмотической обработки. Объединенные данные из n = 11 срезов.
Объемные изменения
На основании данных о морфометрических изменениях толщины среза и расширения границ мы далее попытались оценить объемные изменения, вызванные OGD.Мы сделали несколько предположений: (i) горизонтальное расширение изотропно, и поэтому изменение поверхности среза пропорционально квадрату изменения расстояния между границами среза; (ii) вертикальные сдвиги кортикальной поверхности также изотропны на всем срезе, и поэтому вертикальное набухание на всем срезе равно изменению на уровне L4. Изменения объема среза рассчитывали по следующей формуле:
ΔV = Δ Z + Z0Z0 (Δ X + X0X0) 2 (1), где ΔV — изменение объема среза, нормализованное к контрольному объему, ΔZ — изменение высоты среза, Z 0 — начальная высота, ΔX — изменение ширины среза, X 0 — начальная ширина среза.Результаты этих вычислений представлены на сводных рисунках 6C, D, где были сгруппированы данные о вызванных OGD изменениях LFP, прозрачности среза, ширине среза (рассчитанной по смещению границ) и высоте (от ручной отдельной ячейки). и автоматические сдвиги поверхности срезов объединяются вместе), и объем срезов объединяется вместе с SD, взятым в качестве опорной точки времени. Этот график показывает, что объем среза начинает увеличиваться почти сразу и линейно после SDD со скоростью ~ 1% в минуту до достижения 30 минут после SDD, 130 ± 10% (по оценкам n, = 21 срез для сдвига границ и n. = 4 среза для сдвигов поверхности среза) объема до OGD.
Рисунок 6 . Развитие индуцированного OGD отека ткани среза по отношению к ASD. (A – D) Сводный график собирает временной ход: (A) OGD-индуцированных изменений LFP (среднее значение n, = 54 среза) , (B) прозрачность среза (среднее значение n = 49 срезов), (C) размер среза ( n = 21 срез для сдвига границ и n = 4 среза для сдвига поверхности среза с методом автофокусировки) и (D) объем среза , оцененный из расширение границ среза и возвышение поверхности среза.Все дорожки относятся к электрическому ASD.
Обсуждение
В настоящем исследовании мы охарактеризовали динамику движения массы ткани и объемных изменений, происходящих в срезах коры ствола крысы в условиях OGD. Наши основные результаты заключаются в том, что OGD эффективно вызывает движение массы и набухание тканей, которые характерны для ишемического отека головного мозга, что начало набухания ткани тесно связано с aSD и что отек кортикального среза развивается в относительно медленном временном масштабе после aSD.
Наши данные о развитии индуцированного OGD отека ткани в кортикальных срезах согласуются с результатами предыдущих исследований, в которых увеличение объема среза на ~ 20% наблюдалось через 30 минут после сочетания отравления цианидом с агликемией (Elkin et al., 2010) (настоящее OGD-исследование: на 30%). Это также согласуется с увеличением содержания воды в срезах, которое наблюдалось через 30 минут после метаболического нарушения (MacGregor et al., 2003; Kumar et al., 2006; Elkin et al., 2010). Примечательно, что окончательное увеличение объема срезов достигло ~ 75% к 24 часам после начала метаболического инсульта (Elkin et al., 2010). Следовательно, вероятно, что вызванное OGD набухание среза, которое отслеживалось здесь на начальных этапах формирования отека, также продолжается после 30-минутного интервала. Основная новизна настоящего исследования состоит в том, что показано, что возникновение отека, вызванного OGD, тесно связано с aSD. Ранее развитие цитотоксического отека, проявлявшееся набуханием сомы нейрональных и глиальных клеток, дендритными бусами и сокращением внеклеточного пространства, также проявлялось через несколько минут после аСД (Vanharreveld, 1957; Murphy et al., 2008; Ришер и др., 2009, 2010, 2012). Принимая во внимание эти предыдущие результаты, наши результаты показывают, что цитотоксический отек и отек тканей в кортикальных срезах развиваются параллельно, при этом aSD является их основным триггером. Мы предполагаем, что цитотоксический отек является основной причиной отека тканей в экспериментальных условиях погруженного среза мозга, и что развитие цитотоксического отека поддерживается постоянным и неограниченным буферизацией внеклеточного пространства электролитами и водой, содержащимися в среде ванны.Это может отличаться от настроек in vivo , где доступность электролитов и воды во внеклеточном пространстве ограничена недостатком кровоснабжения в ишемическом ядре в случае фокальной ишемии или во время глобальной ишемии мозга, вызванной остановкой сердца. В последних случаях может развиться цитотоксический отек без выраженного отека тканей. Однако доступность ионов и воды через остаточное / побочное кровоснабжение, а также за счет пассивной горизонтальной диффузии может обеспечить субстрат для чрезмерного цитотоксического действия и набухания тканей, что может привести к более сильному повреждению клеток и еще больше усугубить ишемическое состояние.
Раннее начало набухания ткани после индуцированного OGD aSD может указывать на особую тяжесть этого типа метаболического инсульта в условиях погружения по сравнению с условиями границы раздела или ситуацией in vivo . При приготовлении погруженного среза, используемом в настоящем исследовании, «точка фиксации» необратимого повреждения достигается во время ASD, о чем свидетельствует полное и необратимое устранение синаптического ответа и мембранного потенциала во время эпизодов OGD с ASD, в то время как более короткие эпизоды OGD без ASD показывают функциональное восстановление (Rader, Lanthorn, 1989; Tanaka et al., 1997; Джоши и Эндрю, 2001; Юзекаева и др., 2017). Следовательно, aSD в погруженных срезах является конечным событием, отмечающим гибель нейронов. Напротив, в срезах мозга с границей раздела газ-жидкость, где буферизация внеклеточного пространства ограничена (Croning and Haddad, 1998), SDA не является конечным событием, и срезы могут быть спасены реперфузией с помощью ACSF, содержащего кислород / глюкозу. через несколько минут после индуцированного гипоксией аСД (еще в нормальных гликемических условиях) (Balestrino and Somjen, 1986; Young et al., 1991; Somjen, 2001).Во время глобальной ишемии in vivo aSD также не является конечным событием, поскольку морфологические и функциональные нарушения корковых нейронов обратимы после начала реперфузии в течение ~ 5 минут после aSD (Murphy et al., 2008). Более того, корковые срезы, полученные через 5-6 часов после остановки сердца, обладают вполне нормальными физиологическими и морфологическими свойствами (Charpak, Audinat, 1998). Таким образом, индуцированный OGD aSD в погруженных срезах оказывается особенно разрушительным, что может отражаться ранним началом набухания ткани в связи с aSD.
В заключение, мы показали, что движение массы ткани головного мозга, деформация и набухание также происходит в условиях, подобных ишемии, в кортикальном срезе in vitro , что его начало тесно связано с SD и развивается относительно медленно. после aSD. Очевидно, несколько важных элементов ишемического отека отсутствуют в восстановленной системе среза мозга in vitro , особенно вазогенные механизмы, механические ограничения, вызывающие повышение внутричерепного давления, и компонент набухания, вторичный по отношению к дисфункции глимфатической системы.Следовательно, эта модель далеко не полностью способна воспроизвести ишемический отек in vivo . Однако эта модель достаточно хорошо воспроизводит движение массы мозговой ткани и увеличение объема мозговой ткани, которые происходят во время ишемического отека мозга in vivo . Более того, мы обнаружили, что отек срезов, вызванный OGD, можно уменьшить с помощью гиперосмотического лечения, которое является одним из наиболее часто используемых методов лечения ишемического отека мозга. Таким образом, мы предполагаем, что эта модель может быть полезна для исследования клеточных и молекулярных механизмов, включая специфические ионные каналы, переносчики и другие белки, участвующие в дезадаптивном переносе ионов и воды через нейрональные и астроцитарные мембраны во время ишемического отека, а также при поиске для более эффективного лечения этого разрушительного состояния.
Заявление о доступности данных
Исходные и обработанные данные, а также процедуры обработки и анализа сигналов доступны по запросу у авторов.
Авторские взносы
РК задумал проект. EJ, AG и MM проводили эксперименты. AG и EJ проанализировали данные. РК написал газету.
Финансирование
Работа поддержана РНФ № 17-15-01271 (электрофизиология и визуализация) и субсидией, выделенной Казанскому федеральному университету для выполнения государственного задания в сфере научной деятельности №6.2313.2017 / 4.6 (разработка системы автофокуса) и выполнено в рамках Программы повышения конкурентоспособности Казанского университета.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы благодарим докторов наук. Сергея Кирова и Игоря Медина за полезные предложения и комментарии к рукописи, а также Дэвида Джаппи за редактирование рукописи.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fncel.2018.00502/full#supplementary-material
Список литературы
Эндрю Р.Д., Лаброн М.В., Бёнке С.Э., Карндафф Л. и Киров С.А. (2007). Физиологические доказательства того, что пирамидные нейроны лишены функциональных водных каналов. Cereb. Cortex 17, 787–802. DOI: 10.1093 / cercor / bhk032
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Балестрино, м., и Somjen, G.G. (1986). Хлорпромазин защищает ткань мозга при гипоксии, задерживая распространение опосредованного депрессией притока кальция. Мозг Рес . 385, 219–226. DOI: 10.1016 / 0006-8993 (86)
-XPubMed Аннотация | CrossRef Полный текст | Google Scholar
Бриссон, С.Д., Эндрю, Р. (2012). Популяция нейронов в гипоталамусе, которая значительно сопротивляется острому ишемическому повреждению по сравнению с неокортексом. J. Neurophysiol. 108, 419–430. DOI: 10,1152 / jn.00090.2012
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чарпак С., Аудинат Э. (1998). Остановка сердца у грызунов: максимальная продолжительность совместима с восстановлением активности нейронов. Proc Natl Acad Sci U.S.A. 95, 4748–4753. DOI: 10.1073 / pnas.95.8.4748
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кронинг, М. Д., и Хаддад, Г. Г. (1998). Сравнение конструкций камер для срезов головного мозга для исследования кислородного голодания in vitro . J. Neurosci. Методы 81, 103–111. DOI: 10.1016 / S0165-0270 (98) 00023-5
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Dostovic, Z., Dostovic, E., Smajlovic, D., Ibrahimagic, O.C, and Avdic, L. (2016). Отек мозга после ишемического инсульта. Med. Arch. 70, 339–341. DOI: 10.5455 / medarh.2016.70.339-341
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Драйер, Дж. П., Лемале, К. Л., Кола, В., Фридман, А., и Шокнехт, К.(2018). Распространение деполяризации — это не эпифеномен, а основной механизм цитотоксического отека в различных структурах серого вещества мозга во время инсульта. Нейрофармакология 134, 189–207. DOI: 10.1016 / j.neuropharm.2017.09.027
CrossRef Полный текст | Google Scholar
Дюге, Г. П., Дюмулен, А., Триллер, А., и Дьедон, С. (2005). Целенаправленное использование совместно высвобождаемых тормозных передатчиков в центральных синапсах. J. Neurosci. 25, 6490–6498.DOI: 10.1523 / JNEUROSCI.1500-05.2005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джала В., Халилов И., Бен Ари Ю. и Хазипов Р. (2001). Нейронные механизмы вызванных аноксией сетевых колебаний в гиппокампе крысы in vitro . J. Physiol. 536, 521–531. DOI: 10.1111 / j.1469-7793.2001.0521c.xd
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Элкин, Б.С., Шайк, М.А., и Моррисон, Б.(2010). Фиксированный отрицательный заряд и эффект Доннана: описание движущих сил, связанных с набуханием и отеком мозговой ткани. Philos. Пер. Математика. Phys. Англ. Sci. 368, 585–603. DOI: 10.1098 / rsta.2009.0223
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джоши И. и Эндрю Р. Д. (2001). Визуализация аноксической деполяризации во время ишемических состояний в срезе полушария мозга мыши. J. Neurophysiol. 85, 414–424. DOI: 10.1152 / январь 2001.85.1.414
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джузекаева Е., Насретдинов А., Гайнутдинов А., Синцов М., Мухтаров М., Хазипов Р. (2017). Преимущественное инициирование и распространение аноксической деполяризации в слое 4 коры ствола крысы. Фронт. Cell Neurosci. 11: 390 DOI: 10.3389 / fncel.2017.00390
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хазипов Р., Зайнутдинова Д., Огиевецкий Е., Валеева Г., Митрухина О., Манент Дж. Б. и др. (2015). Атлас постнатального мозга крысы в стереотаксических координатах. Фронт. Нейроанат. 9: 161. DOI: 10.3389 / fnana.2015.00161
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кумар В., Найк Р. С., Хиллер М. и Кляйн Дж. (2006). Эффекты модуляторов потока хлоридов в модели in vitro формирования отека мозга. Brain Res. 1122, 222–229. DOI: 10.1016 / j.brainres.2006.09.012
PubMed Аннотация | CrossRef Полный текст | Google Scholar
МакГрегор, Д. Г., Авшалумов, М. В., и Райс, М. Е. (2003). Отек мозга, вызванный ишемией in vitro : причинные факторы и нейрозащита. J. Neurochem. 85, 1402–1411. DOI: 10.1046 / j.1471-4159.2003.01772.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мерфи Т. Х., Ли П., Беттс К. и Лю Р. (2008). Двухфотонная визуализация начала инсульта in vivo показывает, что независимая от NMDA-рецептора ишемическая деполяризация является основной причиной быстрого обратимого повреждения дендритов и шипов. J. Neurosci. 28, 1756–1772. DOI: 10.1523 / JNEUROSCI.5128-07.2008
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Обейдат А.С., Джарвис К.Р. и Эндрю Р.Д. (2000). Глутамат не опосредует острое повреждение нейронов после распространения депрессии, вызванной недостатком O-2 / глюкозы в срезе гиппокампа. J. Cereb. Метаболизм кровотока. 20, 412–422. DOI: 10.1097 / 00004647-200002000-00024
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Райдер Р.К. и Ланторн Т. Х. (1989). Экспериментальная ишемия вызывает стойкую деполяризацию, блокируемую пониженным содержанием кальция и антагонистами NMDA. Neurosci. Lett. 99, 125–130. DOI: 10.1016 / 0304-3940 (89) -0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ришер В., Ард Д., Юань Дж. И Киров С. А. (2010). Рецидивирующие спонтанные распространяющиеся деполяризации способствуют острому повреждению дендритов в ишемической полутени. J. Neurosci. 30, 9859–9868.DOI: 10.1523 / JNEUROSCI.1917-10.2010
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ришер, В., Крум, Д., Киров, С.А. (2012). Стойкий отек астроглии сопровождает быстрое обратимое повреждение дендритов во время распространяющейся деполяризации, вызванной инсультом. Глия 60, 1709–1720. DOI: 10.1002 / glia.22390
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ришер, В.С., Эндрю, Р.Д., Киров, С.А. (2009). Пассивный объемный ответ астроцитов в реальном времени на острый осмотический и ишемический стресс в кортикальных срезах и in vivo, выявлен с помощью двухфотонной микроскопии. Glia 57, 207–221. DOI: 10.1002 / glia.20747
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шваб С., Ашофф А., Шпрангер М., Альберт Ф. и Хаке В. (1996). Значение мониторинга внутричерепного давления при остром полушарном инсульте. Неврология 47, 393–398. DOI: 10.1212 / WNL.47.2.393
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Танака, Э., Ямамото, С., Кудо, Ю., Михара, С., и Хигаши, Х.(1997). Механизмы, лежащие в основе быстрой деполяризации, вызываемой лишением кислорода и глюкозы в нейронах CA1 гиппокампа крысы in vitro . J. Neurophysiol. 78, 891–902. DOI: 10.1152 / jn.1997.78.2.891
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ванхарревельд А. (1957). Изменение объема нейрональных элементов коры при асфиксии. Am. J. Physiol. 191, 233–242. DOI: 10.1152 / ajplegacy.1957.191.2.233
CrossRef Полный текст | Google Scholar
Янг, Дж.Н., Эйткен П. Г. и Сомьен Г. Г. (1991). Кальций, магний и долгосрочное восстановление после гипоксии в срезах ткани гиппокампа. Brain Res. 548, 343–345. DOI: 10.1016 / 0006-8993 (91)
-R
PubMed Аннотация | CrossRef Полный текст | Google Scholar
SN&R • Реальное дно барреля занятости?
«Здравствуйте! Хочешь пиццы? »
Женщина, покачиваясь, останавливает свою тележку и изучает меня.Ее взгляд начинается с моей головы — бейсболки с логотипом продукта, улыбающимся лицом, именным значком — и она лежит на моем столе, уставленном купонами и дымящимися ломтиками сицилийских пирогов на вынос.
Я могу сказать, что она сидит на диете. Бесплатные образцы — ее заклятые враги. «Я не хочу пиццу!» — объявляет она, победно улыбаясь.
«ОК! Хорошего дня!» Я чирикаю.
Я сканирую проход в супермаркете в поисках новых покупателей, когда она наклоняется и усмехается: «Ну, а как тебе твоя работа?»
Очевидно, что она считает продвижение по службе в супермаркетах дном, и не только она.Однажды я ехал полчаса, чтобы поработать в магазине газировки в Розвилле, все время слушая утренний радиокомик Адам Каролла о жалком характере моей работы. «Это на шаг выше тех, кто стоит на углу и машет знаками!» — сказал он, окружение хихикало на заднем плане.
Итак, когда эта покупательница спрашивает, как мне нравится моя работа, она отвечает, что у нее есть преимущество — в потреблении пиццы и в жизни. Я пожимаю плечами и признаюсь: «Я люблю свою работу».
Она закатывает глаза и толкает тележку к отделению с йогуртами.Она мне не верит.
Может быть, и вы тоже, но посмотрите на мой рабочий день: мне платят за то, что я стою в кондиционированном магазине и слушаю Hall & Oates и Earth, Wind & Fire по системе громкой связи. Я никем не управляю, и меня никто не контролирует. Я купил 12 пицц на чужие деньги и удивляю людей бесплатными кусочками. Многие рады меня видеть, рады получить что-то даром.
Честно говоря, я работал намного больше за гораздо меньшие деньги. Заработок варьируется, но я часто зарабатываю в час на продвижении в супермаркете больше, чем на любой другой работе, которую я имел (включая редактор SN&R по искусству и развлечениям).Люди используют работу по продвижению, чтобы поддержать актерскую карьеру, написание проектов и стартапов. Заработок хороший, график гибкий, а остатки еды заполняют вашу кухню между зарплатами.
Конечно, стоять на публике, взаимодействуя со всеми, кто проходит, есть свои опасности. Один покупатель неоднократно подкрадывался ко мне сзади, чтобы «проверить мои рефлексы». Жуткие мужчины предлагают согреть меня, когда я дрожу в проходе с мороженым. Я слышал о всех диетических ограничениях. Газ, диабет, несварение желудка — для девушки по купону нет ничего личного.
Я разговорный заложник, но иногда это отличный разговор. Однажды африканец прошел мимо незнакомого овоща. Этот экзотический клубень часто был единственным, что его семья ела, когда он рос. Тогда ему это не нравилось, но теперь он не может насытиться.
Его дети не тронут его. «Они даже убирают овощи из пиццы!» он сказал. После десятилетия, проведенного в Америке, его все еще озадачивает концепция придирчивого едока. Это так же странно, как если бы кому-то в магазине, переполненном едой, платили за то, чтобы раздать ее.
Коктейли в бочках
Коктейли в бочках
К настоящему времени мы все знаем, что Busted Barrel — лучший ром, сделанный с любовью здесь, в штате Гарден. Вот несколько отличных коктейлей, которые сделают ваш опыт Busted Barrel еще лучше! Ваше здоровье!!
Девушка из Джерси Мохито
1 выстрел Busted Barrel Silver Rum
3 листа мяты или базилика
3-5 нарезанных ягод клубники (в зависимости от размера)
8 Черника
1/2 ч.л. сахара-сырца
сок 1/2 лайма
клубная газировка
Смешайте клубнику и чернику.Добавьте серебряный ром Busted Barrel, листья мяты или базилика, сахар и сок лайма. Путаница снова, пока ненадолго. Вылейте лед в высокий стакан Collins и сверху полейте содовой.
Украсить долькой лайма.
Джерси Summer Smash
1 выстрел Busted Barrel Silver Rum
1 рюмка Blue J Сиропы Бузины и лимона
6 ежевики
Имбирный эль (или имбирный эль со вкусом ежевики, если вы его найдете)
Смешайте серебряный ром в треснувшей бочке, сироп Blue J и ежевику на дне рок-бокала.Добавьте лед и имбирный эль и хорошо перемешайте. Украсить долькой лимона.
Оборван в Бразилии
2 выстрела Busted Barrel Silver Rum
Сок 1 крупного лайма
1 чайная ложка сырого сверхтонкого сахара
Клубная газировка
В каменной кувшине (или подобном сосуде) смешайте серебряный ром Busted Barrel Silver Rum с соком лайма и сахаром. Сверху полейте содовой и украсьте ломтиком лайма.
Вишня запеченная
2 выстрела Busted Barrel Dark Rum
1 рюмка вишневого бренди
1 бутылка мексиканской колы (хорошего сорта с сахаром, а не кукурузного сиропа)
5 вишен (без косточек)
1/2 гренадина
Осторожно разбейте вишни о дно стакана Tall Collins.добавить в стакан лед. Вылейте в стакан все остальные ингредиенты. Перемешайте, пока хорошо не перемешается. Украсить свежей вишней или мараскино.
Джерси Фреш
2 порции рома в бочках (серебристый или темный, в зависимости от того, что вы предпочитаете)
2 порции сиропов Blue J (любой вкус, все они восхитительны)
Клубная газировка
Наполните бокал льдом. Добавьте ром в бочках и сиропы Blue J и перемешайте. Сверху полейте содовой, размешайте в последний раз и наслаждайтесь!
Это также отличный, легкий удар для больших групп.Наполните чашу для пунша льдом. Налейте 1 бутылку рома Busted Barrel Rum и 2 бутылки сиропа Blue J. Добавьте 1-2 литра содовой.
Разоренная колада
3 выстрела Busted Barrel Silver Rum
2 столовые ложки кокосовых сливок
1/4 стакана свежего ананаса
1 банан
8 унций ананасового сока (желательно свежего)
1 столовая ложка тертого кокоса
Заполните блендер на 3/4 льдом (если вам нравится, сделайте свой собственный лед из кокосовой воды).Добавьте все ингредиенты и взбивайте до однородной массы. Украсить вишней, долькой лайма и измельченным кокосом.
Обезболивающая система Cocktail Whisperer # 201
(по мотивам книги «Аптекарские коктейли» Уоррена Боброу)
Темный ром в бочке, 3 унции
Серебряный ром в бочонке 1 унция
1 ч. Фруктового сиропа
кокосовое молоко 1/2 унции
2 унции свежего ананасового сока
2 унции свежевыжатого апельсинового сока
2 унции свежевыжатого грейпфрутового сока
кубика льда, полученного путем замораживания кокосовой воды
апельсиновый биттер, 4 черточки
тертый мускатный орех
Наполните шейкер Boston на 3/4 простым льдом.Смешайте все жидкие ингредиенты со льдом и энергично встряхивайте в течение 20 секунд. Наполовину наполните стакан колотым льдом, сделанным из кубиков кокосового льда, и процедите смесь в стакан. Украсить копьем ананаса и щепоткой свежего мускатного ореха. Глоток, пока не исчезнет боль!
Низ ствола? Используйте свой садовый урожай
Кэти Вилкенс, MS, RD
Осенью меня всегда радует изобилие, которое возникает в результате небольшой работы, солнца и воды.В прошлом году мой сад произвел 5-галлонный контейнер для тыквы, вероятно, 300 яблок и целое ведро картофеля. Я был так взволнован.
Сейчас весна, и я немного устал от кабачков, яблок и картофеля. У меня не было вдохновения прикончить их, пока я не решил использовать другие методы приготовления, чтобы все изменить. В прошлые выходные я отказался от барбекю и приготовил тыкву на гриле, чтобы она стала привлекательным, интересным и не содержащим соли дополнением к ужину. Я нарезал и обжарил яблоки и поджарил картофель в духовке вместе с цветной капустой.
Мои садовые ведра теперь пусты и чисты, ожидая наполнения весенними продуктами, такими как ревень, капуста, мангольд и салат.
Если у вас нет сада, поищите зимние тыквы, яблоки и картофель на местном рынке.
Приготовленные на гриле кабачки Winter’s End1-2 желудя или тыквы, нарезанных ломтиками толщиной в 2,5 см, без семян.
1-2 столовые ложки оливкового масла
1-2 столовые ложки коричневого сахара
1-2 столовые ложки сливочного масла
Нагрейте гриль, пока он не станет достаточно горячим (около 400 градусов по Фаренгейту).Смажьте кабачки легким слоем оливкового масла, поставьте на гриль примерно на 5 минут и переверните. Когда вилка станет мягкой, смажьте растопленным маслом и коричневым сахаром. Оставьте на гриле на 1 минуту, выньте и подавайте.
Информация о питании: (на 1 порцию чашки или 2-3 раунда):
калорий: 99, углеводов: 19 грамм, белков: 1 грамм, натрия: 6 миллиграммов
Жареные яблоки с грецкими орехами3-4 яблока, нарезанные ломтиками
1-2 столовые ложки сливочного масла
½ чайной ложки корицы
¼ чайная ложка мускатного ореха
¼ чайная ложка кардамона
½ стакана грецких орехов
Половина лимона
Растопить масло на сковороде и добавить яблоки и специи.Перемешайте, пока не станет мягким. Сверху посыпать орехами и варить еще 1-2 минуты. Снять с огня. Выжмите сок из половины лимона и подавайте.
Информация о пищевой ценности:
калорий: 218, углеводов: 28 г, белков: 3 г, натрия: 3 миллиграммов
Жареный картофель в духовке3-5 картофеля
2 столовые ложки оливкового масла
2 столовые ложки тертого сыра Пармезан
Установить духовку на 500 градусов F.Нарежьте картофель на квадраты размером около 2,5 см и выложите на противень. Сбрызнуть маслом, а затем перемешать. Поставить в духовку и выпекать 8-10 минут. Перемешайте и готовьте еще 8-10 минут. Протестируйте вилкой. Когда все будет готово, достаньте из духовки и посыпьте 2 столовыми ложками натертого на терке пармезана. Подавать немедленно.
Информация о пищевой ценности:
калорий: 191, углеводов: 28 г, белков: 4 г, натрия: 58 г
Информация в этой колонке предназначена для людей, которые хотят сохранить здоровье почек и снизить артериальное давление, соблюдая диету с низким содержанием натрия.В большинстве случаев, за исключением диализных пациентов, считается, что диета с высоким содержанием калия помогает снизить высокое кровяное давление. Эти рецепты не предназначены для людей, находящихся на диализе, без наблюдения диетолога.
[Кэти Г. Уилкенс — зарегистрированный диетолог и руководитель отделения Северо-Западного почечного центра. Получив награду Сьюзан Кнапп за выдающиеся достижения в области образования от Совета Национального фонда почек по почечному питанию, она имеет степень магистра наук в области диетологии Вашингтонского университета.См. Другие ее рецепты на сайте www.nwkidney.org .]
Нацеливание на шиповидные звездчатые клетки стволового поля с использованием везикулярного моноаминергического транспортера 2-Cre мышей линии
Животные
Процедуры для животных были одобрены местным этическим комитетом в Упсале (Uppsala djurförsöksetiska nämnd) и следовали Директиве 2010/63 / EU Европейского парламента и Совета, Шведскому закону о защите животных (Djurskyddslagen: SFS 1988: 534), Шведскому постановлению о защите животных (Djurskyddsförordningen: SFS 1988: 539), а также положения об использовании животных в научных целях: DFS 2004: 15 и SJVFS 2012: 26.Линия Vmat2-Cre (STOCK Tg (Slc18a2-cre) OZ14Gsat / Mmucd (MMRRC)) использовалась отдельно или скрещивалась с репортерной линией tdTomato (Gt (ROSA) 26Sor tm14 (CAG-tdTomato) Hze ; Allen Brain Институт). Следующие праймеры были использованы для идентификации Cre и tdTomato соответственно; Cre 5′-acgagtgatgaggttcgcaaga-3 ‘(прямой, мутантный аллель), 5′-accgacgatgaagcatgtttag-3’ (обратный, мутантный аллель) tdTomato 5′-tgttcctgtacggcatgg-3 ‘(прямой, мутантный аллельccatgta’-gtagtag-gtagtag-3′ (прямой, мутантный аллельcccatgta’- 3 ‘(обратный мутантный аллель), 5′-aagggagctgcgtggagta-3′ (прямой аллель дикого типа), 5’-ccgaaaatctgtgggaagtc-3 ‘(обратный аллель дикого типа).Аллель Vmat2-Cre оставался гетерозиготным, и в анализ включались как самцы, так и самцы мышей, если не указано иное. Исследование проводилось в соответствии с рекомендациями ARRIVE.
Визуализация Vmat2-Cre; экспрессия tdTomato
Взрослый (5 самок и 1 самец, 12–16 недель) Vmat2-Cre; Мышей tdTomato перфузировали (см. Ниже), используя PBS, а затем 4% формальдегид (FA). Вырезали весь мозг, спинной мозг и ткань ганглиев задних корешков. Мозг и два спинного мозга выдерживали в 4% FA в течение ночи при 4 ° C, затем помещали в 4% агарозу (VWR) и разрезали на срезы толщиной 60 мкм с использованием вибратома (Leica VT1200).После инкубации FA один спинной мозг с интактной тканью ганглиев дорсальных корешков помещали в 15% сахарозу (Sigma-Aldrich, США) в 1 × PBS на 24 часа с последующей 24-часовой инкубацией в 30% сахарозе в 1 × PBS в течение 24 часов. h для криозащиты. После этого ткань помещали в среду с оптимальной температурой резки (OCT) (Bio-Optica, Милан, Италия) и мгновенно замораживали на сухом льду в изопентане -80 ° C (Sigma-Aldrich, Германия). Ткани разрезали на срезы 18 мкм с использованием криостата (Leica Cryocut 1800) и собирали на предметные стекла Superfrost Plus (Thermo Scientific).
Срезы впоследствии визуализировали с помощью флуоресцентного микроскопа Olympus BX61WI (Olympus, Япония), где изображение красной флуоресценции (выражение tdTomato) отображали и редактировали с помощью программного обеспечения ImageJ (ImageJ, США) и Adobe Photoshop (для создания композитов). Для определения анатомического положения ткани использовались атласы 38,39 .
Микроинъекция вирусных векторов
Для маркировки таламокортикальных проекций в коре бочкообразного поля, взрослый Vmat2-Cre; tdTomato Мышам (5 самок, возраст 10–11 недель) вводили 500 нл AAV1.Вирус hsyn.hChR2 (h234R) -EYFP (сокращенно AAV1.hsyn-ChR2.eYFP в разделе результатов, номер партии v67891) от Addgene в титре 1,1 × 10 13 мкг / мл в области VPM таламуса. Координаты: B: -1,80; ML: 1,75; ДВ: 3,70. В качестве альтернативы, чтобы пометить поле кортикального ствола Vmat2-Cre клеток у взрослых мышей (2 самцов и 3 самки, возраст 11 недель), такой же объем AAV8.hsyn-DIO-mCherry (номер партии v61605, титр: 2,20 × 10 13 cg / ml) от Addgene, или AAVDJ.EF1a-DIO-HTB (дата партии 2017, титр 1.83 × 10 11 вг / мл) вируса, произведенного основным центром Salk Institute GT3 (перенос генов, нацеливание и терапия), вводили в кору S1 мышей Vmat2-Cre (2 самки в возрасте 10–23 недель) . Используемые координаты: B: 0,00; ML: 2,75; ДВ: 0,90.
Препарат ткани для электрофизиологии
Мышей (7 самцов, 8 самок; возраст 12–52 недели) анестезировали ~ 0,5 мл изофлурана (FORANE, Baxter, США) в течение 1–2 минут и вводили внутрибрюшинно 0,3 мл кетамина ( Кеталар, 10 мг / мл, Pfizer) и 0.3 мл Медетомидина (Домитор, 1 мг / мл, Орион Фарма). Примерно через 4–5 минут эффективность анестезии измеряли по рефлексу защемления лапы. Мышь фиксировали на папиросной бумаге с помощью ленты и удаляли кожу над грудной клеткой. Часть грудной клетки была осторожно удалена, чтобы обнажить сердце. Иглу для перфузии вводили в левую брюшную часть и делали небольшой разрез в правом предсердии. Мыши перфузировали холодным режущим раствором на основе NMDG-HEPES (концентрация в мМ: 93 N-метил-d-глюкамин, 2.50 KCl, 1,20 NaH 2 PO 4 , 30 NaHCO 3 , 20 HEPES, 25 глюкоза, 5 аскорбат натрия, 2 тиомочевина, 3 пируват натрия, 10 MgSO 4 ∙ 7H 2 O, 0,5 CaCl 2 ∙ 2H 2 O) примерно на 4 мин с помощью перистальтического насоса со скоростью 450 мл / час. Мозг был осторожно удален из черепа, и коронковые срезы толщиной 300 мкм были сделаны с помощью вибратома (Leica vt1200) в холодном непрерывно насыщенном кислородом (95% O 2 и 5% CO 2 ) режущем растворе (как указано выше) из В: — 0.От 10 до — 1,46 мм по стереотаксическим координатам атласа мозга мыши 38 . Нарезанные срезы затем восстанавливали в aCSF (концентрация в мМ: 126 NaCl, 2,5 KCl, 1,25 NaH 2 PO 4 , 26 NaHCO 3 , 10 глюкозы, 1,5 CaCl 2 , 1,5 MgCl2) при 36 °. C в течение 1 ч и при комнатной температуре не менее 30 мин перед помещением в записывающую камеру.
Электрофизиология
Затем срезы были перенесены в записывающую камеру для электрофизиологического анализа в соответствии с ранее описанной процедурой 40 , где нейроны Vmat2-Cre были идентифицированы с использованием водно-иммерсионного объектива 60х или 20х (LUMPlan FI, 0.90 числовая апертура (NA), Olympus) с помощью красной флуоресценции (tdTomato). Короче говоря, нейроны визуализировались на камере Zyla sCMOS (Andor Technology Ltd), подключенной к зеленому (550 нм, система CoolLED) флуоресцентному светодиодному источнику света. Патч-электроды (6–10 МОм) извлекали из капилляров из боросиликатного стекла (GC150F-10 Harvard Apparatus) с помощью пылающего / коричневого съемника для микропипеток (P-1000, Shutter Intrument, США). Внутренний раствор содержал (в мМ): 130 K-глюконат, 40 HEPES, 1,02 MgCl 2 , 2.17 MgATP, 0,34 NaGTP, pH доведен до 7,2, используя 1 M KOH. Потенциал жидкого соединения корректировали перед каждым патченным нейроном. Сигналы от целых клеток были усилены усилителем MultiClamp 700B (Axon Instruments), оцифрованы с частотой 20 кГц с помощью Digidata 1440A (Molecular Devices), отфильтрованы нижними частотами с частотой 10 кГц и получены с помощью программного обеспечения WinWCP (д-р Дж. Демпстер, Университет Стратклайда, Глазго, Великобритания). Анализ данных проводился с помощью Clampfit 10.3 (Molecular Devices, США), Mini Analysis (Synaptosoft, США) и Matlab (Mathworks).Частота срабатывания потенциала действия рассчитывалась путем инвертирования интервала между всплесками (I-S-I), который представляет собой временной интервал между двумя соседними пиками потенциала действия.
Для проверки синаптической связи между полем ствола кортикального Vmat2-Cre; tdTomato нейронов и таламокортикальных афферентных проекций, экспрессия канала родопсина 2 (ChR2) в области VPM таламуса была впервые подтверждена под 20-кратным водно-иммерсионным объективом (LUMPlan FI, числовая апертура (NA) 0,90, Olympus) с помощью желтого флуоресценция (eYFP).В оптогенетическом эксперименте область, экспрессирующую ChR2, стимулировали синим светом (473 нм) с использованием (Mic-LED, Prizmatix, Израиль) через оптическое волокно, размещенное примерно в 5 мм от области под углом 30 градусов при частоте 1 Гц (10 мс длительность импульса при 6 мВ / мм ( 2 световой мощности), в то время как ответы от нейронов tdTomato измеряли как ВПСП в конфигурации токовых клещей для всей клетки. Задержка была определена как временная задержка от начальной точки светового импульса до активации соответствующего EPSP.
Окрашивание нейробиотином и визуализация
Нейробиотин Tracer (Vector Laboratories, США) добавляли во внутриклеточный раствор (2–4 мг / мл) и доставляли в клетку-мишень во время записи патч-кламп. Деполяризующие импульсы тока (0,5–1 нА, длительность 150 мс) вводились в клетку через регистрирующий электрод с частотой 3 Гц в течение 5–10 мин для улучшения диффузии нейробиотина в нейритах. После записи патч-пипетка осторожно вынималась из ячейки и удалялась из среза.Срез дополнительно перфузировали в течение не менее 15 минут для удаления избыточного нейробиотина в ткани, который затем фиксировали в 4% параформальдегиде (PFA, Histolab) в течение ночи при 4 ° C. Фиксированные срезы промывали фосфатно-солевым буфером (PBS, Fisher BioReagents) трижды по 10 минут каждый и инкубировали в окрашивающем растворе, содержащем конъюгат стрептавидина Alexa Flour 488 (Invitrogen), с коэффициентом разбавления 1: 1000 в 0,3%. Раствор Triton X-100 (Sigma) PBS в течение 4 часов при комнатной температуре на качалке, а затем инкубирование при 4 ° C на качалке в течение 72 часов для визуализации тонких процессов.Окрашивающий раствор удаляли после инкубации, и срез промывали 0,3% раствором TritonX PBS два раза по 20 минут каждый, а затем один раз промывали раствором PBS в течение 20 минут. Затем срез помещали на предметное стекло микроскопа с установочной средой, содержащей DAPI (противоуглеродный реагент ProLong Gold с DAPI, Invitrogen, США). Поверх среза осторожно помещали покровное стекло, а края покрывали лаком для ногтей. Изображение закрепленного среза получали с помощью конфокального микроскопа Leica TCS SP8 с погружением в воду 25 × (HCX IRAPOL, 0.95 NA) объективный. Изображенные клетки были отслежены и реконструированы с использованием плагина Simple Neurite Tracer в программном обеспечении NIH ImageJ (Национальные институты здравоохранения, Бетесда, Мэриленд). Наличие дендритных шипов использовалось для различения аксонов и дендритов, и анализ был сосредоточен на дендритных процессах, чтобы избежать неполных реконструкций потенциально разрезанных аксонов.
Диссоциация ткани для секвенирования одной клетки
Две взрослых (48–55 недель) самки Vmat2-Cre; tdTomato и одна tdTomato отрицательная самка (возраст 11 недель) были подвергнуты седации в 4% изофлурансодержащем боксе (FORANE, Baxter, США) и инъецированы i.p с 0,6 мл (1/1) смеси кетамина (Ketalar, 10 мг / мл, Pfizer) и Медетомидина (Domitor, 1 мг / мл, Orion Pharma). После этого животных перфузировали ледяной предварительно оксигенированной 1 × PBS Дульбекко (DPBS, Sigma-Aldrich). В том же растворе передний мозг был разрезан на срезы 400 мкм с помощью вибратома (Leica VT1200). Поскольку многие клетки tdTomato можно было обнаружить в полосатом теле, кортикальные области tdTomato и эквивалентные области контрольной ткани были осторожно изолированы с помощью ножниц.После этого с корковой областью каждого животного обрабатывали отдельно для диссоциации. Диссоциацию проводили с использованием набора для диссоциации мозга взрослых с небольшими изменениями инструкций производителя (Miltenyi Biotec, номер по каталогу: 130-107-667). Срезы помещали в пробирки Эппендорфа, содержащие предварительно нагретый (37 ° C) фермент 1 (475 мкл буфера Z, 25 мкл фермента P), куда добавляли 15 мкл предварительно нагретой смеси ферментов 2 (10 мкл буфера Y, 5 мкл фермента A). . Ткани диссоциировали в течение 30 мин при 37 ° C; каждые 10 мин ткани растирали 10 раз с использованием стеклянных пипеток Пастера, диаметр которых уменьшался с каждым сеансом растирания.Для инактивации ферментов растертые суспензии клеток добавляли к 5 мл охлажденного льдом DPBS. Интеллектуальный фильтр (40 мкм, Corning) предварительно замачивали в 3 мл DPBS, после чего суспензию клеток добавляли к фильтру и дополнительно добавляли 2 мл DPBS к фильтру для сбора клеток, возможно оставшихся в фильтре. Образцы центрифугировали в течение 10 минут при 300 g (4 ° C), после чего супернатанты были удалены и осадки ресуспендировали в 450 мкл буферного раствора (0,5% BSA в DPBS) и 50 мкл шариков для удаления миелина (Miltenyi Biotec, номер по каталогу: 130-096-733).Образцы помещали при 4 ° C на 15 минут и добавляли 5 мл буферного раствора перед 10-минутным центрифугированием при 300 g (4 ° C). Супернатанты отбрасывали, а осадки ресуспендировали в 500 мкл буферного раствора, а колонку LS (Miltenyi Biotec, номер по каталогу: 130-042-401) предварительно замачивали в 3 мл буферного раствора. Суспензию клеток добавляли в колонку LS и в колонку добавляли 2 × 500 мкл буферного раствора для сбора клеток, оставшихся в колонке. Суспензии клеток хранили на льду до сортировки, которую проводили сразу после диссоциации.
Сортировка клеток с активацией флуоресценции
Клеточные суспензии были проанализированы, и 178 Vmat2-Cre; tdTomato Клетки сортировали на 384-луночном планшете Smart-Seq2 с использованием программного обеспечения BD FACSMelody и BD FACSChorus. Первоначально стробирование частиц производилось на основе площади прямого и бокового рассеяния света (FSC-A / SSC-A) для обнаружения однородных по характеру частиц размером с клетку, которые затем регулировались по высоте и ширине в два этапа (1. SSC- H / SSC-W, 2. FSC-H / FSC-W) для выделения отдельных клеток.Чтобы обнаружить tdTomato положительных клеток и исключить одноклеточные частицы с автофлуоресценцией, одиночные клетки были заблокированы по интенсивности различных настроек красного и дальнего красного каналов (PE-Cy5 (YG) -A, PE-Cy7 ( YG) -A, (YG) -A) (Рисунок S6). Кортикальный образец от tdTomato отрицательного животного использовался в качестве контрольного образца для определения базовой интенсивности красного и дальнего красного каналов (Рисунок S6A), где после tdTomato порог интенсивности мог быть обнаружен в (YG) -A. канал (рисунок S6B).После этого положительные единичные клетки tdTomato сортировали на планшете, который хранили при -80 ° C до секвенирования библиотеки.
Библиотека секвенирования
Данные для отдельных ячеек были созданы с использованием метода Smart-Seq2 20 . Чтобы нормализовать глубину секвенирования и длину гена, значения экспрессии рассчитывались как считывания на килобазу генной модели и миллион отображаемых считываний (RPKM). После этого вычисляли значения экспрессии для каждого гена 41 с использованием однозначно выровненных считываний и корректировки на однозначно выравниваемые положения с использованием MULTo 42 .Сначала образцы были проанализированы путем демультиплексирования файлов fastq с помощью деиндексатора (https://github.com/ws6/deindexer) с использованием адаптеров индекса nextera и макета 384. После этого отдельные файлы fastq были сопоставлены с соответствующей сборкой генома с использованием выравнивателя STAR 43 с использованием двухпроходного выравнивания для повышения производительности чтения стыков de novo, отфильтрованных только для считывания однозначного сопоставления.
Обработка и кластеризация данных отдельной ячейки
The Vmat2-Cre; Клетки tdTomato секвенировали с использованием Smart-Seq2 20 , где можно было обнаружить в общей сложности 24 582 гена.Данные были проанализированы с помощью SCANPY 44 в Python 3.8.1. Полный код и файл ткацкого станка необработанного файла Vmat2-Cre. tdTomato данные можно найти по адресу: (https://github.com/HannahBanarne/Somatosensory-cortical-Vmat2-analysis). Для базовой предварительной обработки сначала были рассчитаны показатели общей экспрессии генов, ERCC и митохондриальных генов (SCANPY, query.mitochondrial_genes («mmusculus»), pp.calculate_gc_metrics) 45 . Построив распределение полученных показателей (Seaborn, Jointplot), клетки с распределенным количеством генов и широким захватом генов (SCANPY, ‘log1p_n_genes_by_counts’> 6.0, ‘pct_counts_in_top_50_genes’ <80), высокое общее количество ('log1p_total_counts'> 11), низкое количество митохондриальных генов («pct_counts_mito» <0,1) и менее 10% ERCC. Из 178 секвенированных клеток Vmat2-Cre 158 соответствовали критерию включения и были использованы для анализа экспрессии. Для фильтрации генов все гены, которые экспрессировались менее чем в 3 клетках в наборе данных, были исключены, в результате чего 18430 генов были использованы для анализа 158 нейронов Vmat2-Cre (SCANPY, стр.filter_genes). После этого количество отсчетов на ячейку было нормализовано до среднего количества отсчетов (SCANPY, pp.normalize_per_cell) с последующей нормализацией (SCANPY, pp.log1p). Для уменьшения размерности данных был проведен анализ главных компонентов (PCA) (SCANPY, pp.pca) 46 с использованием 1000 наиболее вариабельных генов (SCANPY, pp.highly_variable_genes) 47,48 , а топология данных была рассчитана с использованием Метод UMAP (SCANPY, pp.neighbors ((n_neighbors = 10, metrics = ‘euclidean’, method = ‘umap’, n_pcs = 9), tl.umap)) 35,49 . Чтобы исследовать, существуют ли какие-либо подкластеры нейронов Vmat2-Cre со специфической экспрессией генов, клетки были сгруппированы в подгруппы с использованием метода Лейдена 50 и нанесены на график с помощью UMAP (SCANPY, pl.umap). В результате кластеризации было получено 8 кластеров, и для обнаружения различий в экспрессии генов между кластерами были рассчитаны z-баллы и скорректированные p-значения (Бенджамини-Хохберг для корректировки частоты ложных открытий (FDR)) для верхних дифференциально экспрессируемых (DE) гены в каждом кластере (SCANPY, tl.rank_genes_groups). Экспрессия генов для 10 лучших генов DE из каждого кластера была визуализирована для всех отдельных нейронов с помощью тепловой карты (SCANPY, tl.rank_genes_groups_heatmap) и среднего выражения для этих генов с использованием матричного графика (SCANPY, pl.rank_genes_groups_matrixplot). Для одного из кластеров главные гены DE включали Fcer1g , C1qb и Ctss , которые являются маркерными генами для микроглиальных клеток (mousebrain.org), и, таким образом, 7 клеток, принадлежащих к этому кластеру, были исключены.Уменьшение размеров, топология данных (n_pcs = 9), кластеризация и обнаружение главных генов DE были повторно выполнены для оставшихся 151 нейрона в наборе данных.
Анализ маркерного гена Vmat2-Cre
Анализ экспрессии специфического гена, включая Slc18a2 ( Vmat2 ) и маркерные гены для синтеза серотонина, а также маркеры для возбуждающих и тормозных нейронов, был проведен для общей популяции Vmat2-Cre. (SCANPY, мн. Сложенная_ скрипка). Чтобы исследовать ввод нейротрансмиттера в популяции Vmat2-Cre, «target_and_families.csv ‘был загружен из Guide to Pharmacology (https://www.guidetopharmacology.org), и была рассчитана распространенность экспрессии субъединиц лиганд-зависимых ионных каналов и рецепторов, связанных с G-белком (GPCR) (ген считался выраженным, если log1p = > 0,25), где были визуализированы 50 наиболее распространенных генов соответствующего типа рецептора (SCANPY, pl.stacked_violin).
Прогнозирование типа клеток Vmat2-Cre
Чтобы изучить расположение и молекулярную идентичность популяции Vmat2-Cre, экспрессию маркерных генов кортикального слоя 25 исследовали в популяции Vmat2-Cre.Для дальнейшего изучения кортикальной идентичности нейронов Vmat2-Cre наш набор данных был объединен и выровнен с поднабором кортикальных нейронов из набора данных проектирующих нейронов конечного мозга, опубликованного Zeisel et al. (2018). Набор данных l6_r3_telencephalon_projecting_neurons.loom был получен из (http://www.linnarssonlab.org), который содержит 28 858 отдельных клеток и 27 998 генов, полученных с использованием методов 10-кратной геномики. Из этого набора данных все нейроны, принадлежащие кортикальным пирамидным слоям 2 / 3–6, были выделены для анализа, в результате было получено 9435 клеток и 27 998 генов.Эти клетки были дополнительно обработаны с теми же параметрами, что и набор данных Vmat-Cre, за исключением исключения нейронов с высокой экспрессией ERCC, поскольку метод 10x Genomics не включает ERCC. В результате базовой предварительной обработки было получено 9369 клеток и 17 304 гена. Чтобы увидеть, насколько хорошо два набора данных согласованы друг с другом, нормализованные наборы данных Vmat2-Cre и Zeisel были объединены на основе всех генов из обоих наборов данных (SCANPY, concatenate (join = ‘external’)), а размеры данных были уменьшены и визуализированы с помощью PCA (стр.pca, pl.pca_scatter). После этого из обоих наборов данных были обнаружены верхние 5000 генов с высокой степенью вариабельности (SCANPY, pp.highly_variable_genes), которые в дальнейшем использовались для объединения нормализованных и масштабированных наборов данных об экспрессии общих генов (SCANPY, pp.log1p, pp.scale) и корректировки пакетные эффекты с использованием соответствия взаимных ближайших соседей (MNN) (SCANPY, external.pp.mnn_correct) 35 . Размеры данных были уменьшены с помощью PCA (SCANPY, pp.pca), а топология данных визуализирована с помощью UMAP (pp.neighbors (n_neighbors = 10, n_pcs = 9), pp.umap). Аннотированные вероятные корковые локации и кластеры, оцененные Zeisel et al. были впоследствии предсказаны и назначены клеткам Vmat2-Cre путем первого обучения линейной логистической регрессии со стохастическим градиентным спуском (SGD) обучающего классификатора (scikit-learn, SGDClassifier (loss = ‘log’)) на Zeisel et al. экспрессия гена в качестве переменных-предикторов и аннотированные местоположения или кластеры в качестве целевых меток. После этого были предсказаны вероятные метки местоположения для ячеек Vmat2-Cre, и вероятность правильного присвоения меток была вычислена для каждой ячейки Vmat2-Cre (scikit-learn, pred_proba) перед отображением обратно предсказанных меток и оценок вероятности в масштабированный объединенный набор данных и исходный набор данных Vmat2-Cre.Чтобы визуализировать, насколько хорошо аннотации были предсказаны для ячеек Vmat2, оценки вероятности были нанесены на график распределения (Seaborn, distplot).
Препарат ткани in situ
Две самки взрослых мышей Vmat2-Cre (возраст 10 недель), которым микроинъектировали AAVDJ.EF1a-DIO-HTB в SB1F, перфузировали изофлуран через 14 дней после инъекции (FORANE, Baxter, США) с последующей внутрибрюшинной инъекцией 0,6 мл (1: 1) кетамина (Ketalar, 10 мг / мл, Pfizer) и Медетомидина (Domitor, 1 мг / мл, Orion Pharma).Чтобы свести к минимуму риск заражения и изменения экспрессии генов, животных перфузировали в автоклавированном ледяном 1xPBS. Мозг быстро препарировали и в автоклаве на льду 1xPBS выделяли область полушария, содержащую S1BF, а затем помещали в среду OCT (Bio-Optica, Милан, Италия). Ткани немедленно замораживали на сухом льду в изопентане -80 ° C (Sigma-Aldrich, Германия) и хранили при этой температуре до срезов. Ткани подвергали криосрезу (Leica Cryocut 1800) на срезы толщиной 12 мкм и собирали сериями из 6 слайдов / серию на слайды Superfrost Plus (Thermo Scientific).Чтобы предотвратить деградацию и контаминацию мРНК, завершенные серии хранили при -21 ° C до завершения срезов. Срезы хранили при -80 ° C до начала протокола набора RNAscope Fluorescent Multiplex (Advanced Cell Diagnostics (ACD), каталожный № 320850).
Флуоресцентная гибридизация in situ
Флуоресцентная гибридизация in situ (FISH) была проведена с использованием набора RNAscope Fluorescent Multiplex (Advanced Cell Diagnostics (ACD), номер по каталогу 320850) в соответствии с рекомендациями ACD для свежезамороженных тканей с незначительными модификациями 51 .Вкратце: подлежащие использованию тканевые слайды были взяты при температуре -80 ° C и немедленно зафиксированы в RT 4% PFA в 1 × PBS (Histolab, Швеция) в течение 15 минут, а затем промыты в автоклавированном 1 × PBS в течение 2 минут. После этого ткани дегидратировали с постепенным увеличением концентрации EtOH; 3 минуты в 50%, 3 минуты в 70% и 2 раза по 5 минут в 100% (Merck KGaA, Damstadt, Германия). Слайды помещали при комнатной температуре на 5 минут для высыхания, после чего вокруг интересующей области предметного стекла (2–5 слайдов на зонд и животное) создавали гидрофобный барьер с помощью ручки ImmeEdge (Vector Laboratories, Burlingame, California, USA).После этого срезы инкубировали в протеазе IV в течение 30-40 минут при комнатной температуре с последующей трехкратной 5-минутной промывкой в автоклавированном 1xPBS с последующей инкубацией с зондами-мишенями (каталожный номер: Vglut1 ( Slc17a6 ): 416631-C2, Vglut2 ( Slc17a6 ): 319171-C2; Vglut2 : 319171-C3; Viaat ( Slc32a1 ): 319191-C3; Vmat2 : 42547471-Corb; Scnn1a : 441391-C3, 1:50 в разбавителе зонда, каталожный номер 300041) в течение 2 ч при 40 ° C в печи для гибридизации (HybEZ II Oven, ACD).Следующие этапы амплификации выполняли при 40 ° C в печи, и срезы промывали 2 раза по 2 мин в промывочном буфере RT между каждым этапом амплификации; AMP 1-FL на 30 минут, AMP 2-FL на 15 минут, AMP 3-FL на 30 минут и AMP 4-FL на 15 минут. Стадия окрашивания с использованием AMP 4-FL была выполнена для обеспечения возможности комбинации с вирусным GFP. Наконец, предметные стекла промывали 2 раза по 2 мин в промывочном буфере перед 30-секундной инкубацией в DAPI и помещением в среду для фиксации флуоресценции, предотвращающую исчезновение флуоресценции (Abcam).Предметные стекла покрывали предметными стеклами (Menzel-Gläser) и оставляли для высыхания при 4 ° C. Слайды хранили при этой температуре до визуализации.
Получение и количественная оценка изображений гибридизации in situ
Изображения срезов Vmat2-Cre.HTB SB1F, обработанных РНКскопом, получали с широким полем 20-кратного увеличения с использованием флуоресцентного микроскопа Olympus BX61WI (Olympus, Япония). Канал для каждого датчика был оптимизирован для яркости и контрастности во время получения изображения и количественной оценки.Изображения RNAscope были вручную количественно оценены с использованием плагина Fiji (ImageJ 1.52f) Cell Counter 52,53 . Все клетки Vmat2-Cre.HTB с перекрытием DAPI считались клетками, и одно считывание целевого зонда можно было визуализировать как одну точку. Клетка Vmat2-Cre.HTB считалась экспрессирующей целевой ген, если количество перекрывающихся точек ≥ 3. Результат представлен как процент ± SEM клеток Vmat2-Cre.HTB, экспрессирующих Vmat2 , Vglut1 , Vglut2 , Viaat, Rorb и Scnn1a .
Иммуногистохимия
Один 16-недельный мужчина и две 14-недельные самки Vmat2-Cre; Мышей tdTomato перфузировали PBS, а затем 4% FA. Весь мозг был вырезан и оставлен в 4% FA в течение ночи при 4 ℃. Перед нарезкой ткани помещали в 4% агарозу (VWR) и разрезали на срезы толщиной 60 мкм с использованием вибратома (Leica VT1200). Иммуноокрашивание кроличьими антителами против ГАМК (Sigma-Aldrich) 1: 750 в блокирующем растворе (5% ослиная сыворотка (Sigma-Aldrich), 3% BSA (Sigma-Aldrich) в TBS) (10 срезов на животное).